Location: Home >> Detail
Crop Breed Genet Genom. 2019;1:e190015. https://doi.org/10.20900/cbgg20190015

1 School of Agriculture, Ningxia University, Yingchuan, Ningxia 750021, China
2 Sustainable Perennial Crops Laboratory (SPCL), Beltsville Agricultural Research Center, USDA-ARS, 10300 Baltimore Avenue, Beltsville, MD 20705, USA
* Correspondence: Dapeng Zhang.
This article belongs to the Virtual Special Issue "Genetic Gains in Plant Breeding"
Chinese jujube is one of the earliest domesticated fruit trees in the world. This fruit crop is becoming increasingly popular globally for its outstanding adaptability to marginal land and a broad range of climate conditions. There is a growing demand by growers and consumers for new cultivars with improved yields, earliness, resistance to biotic and abiotic stresses and fruit quality attributes. Breeding jujube for disease resistance is still in its infancy despite the consensus that disease is the top-ranking production constraint and growing disease resistant cultivars is the most sustainable and cost-effective approach for diseases management. Major knowledge gaps remain to be filled in the current breeding scheme for enhancing disease resistance. These knowledge gaps include, but are not limited to (1) a lack of understanding of jujube pathogens in terms of their taxonomy, epidemiology and genetic diversity; (2) a lack of standardized screening methods to identify host resistance; (3) a high rate of germplasm mislabeling; and (4) ineffective production of hybrid progenies. Nonetheless, recent progress in utilization of male sterile parents in controlled crosses, coupled with mounting genomic information on both pathogens and jujube, will likely speed up the combination of host resistance with other horticultural traits. This article reviews the status of jujube disease resistance breeding and suggests future research needs for developing disease resistant cultivars of Chinese jujube. Although the review focuses on jujube breeding programs in China, the summarized information should be useful to researchers in the international jujube community as well.
Chinese jujube (Ziziphus jujuba Mill.) is an economically important fruit crop in the genus Ziziphus Mill. of the Rhamnaceae family. The species is native to China, together with another 13 species in the genus of Ziziphus, with its putative center of origin located in the Yellow River basin in northern China [1,2]. There is another cultivated sister species Ziziphus mauritiana (Indian jujube or Indian ber), which is a tropical and subtropical evergreen species [3]. Although the two cultivated Ziziphus species are in different sections of the Genus Ziziphus (Z. mauritiana in Section Perdurans and Z. jujuba in Section Ziziphus), viable embryos were obtained from Z. mauritiana × Z. jujuba crosses, suggesting that there is inter-specific cross compatibility [4]. Jujube is one of the earliest domesticated fruit trees in China, with a utilization history of more than 7000 years [5–7]. It is believed that current cultivars descend from materials originally selected from sour jujubes (Z. jujuba var. spinosa) which are still widely distributed in north, central and southwest China [5,8]. Chinese Jujube is becoming increasingly popular for its outstanding adaptability, nutritious fruits and multipurpose utilization in food and traditional medicine. It is an ideal crop for arid and semiarid areas of the temperate and subtropical regions where other common fruit trees do not grow well. This feature makes jujube an important economic crop for low income regions and contributes significantly to poverty reduction and reforestation in China (Figure 1). Presently it is ranked as the first dry fruit and the seventh fresh fruit tree in China in terms of cultivation area (600,000 ha), with an annual production of more than 11 million metric tons [6]. The market value of the jujube value chain was estimated about $14–16 billion in 2017 [6].
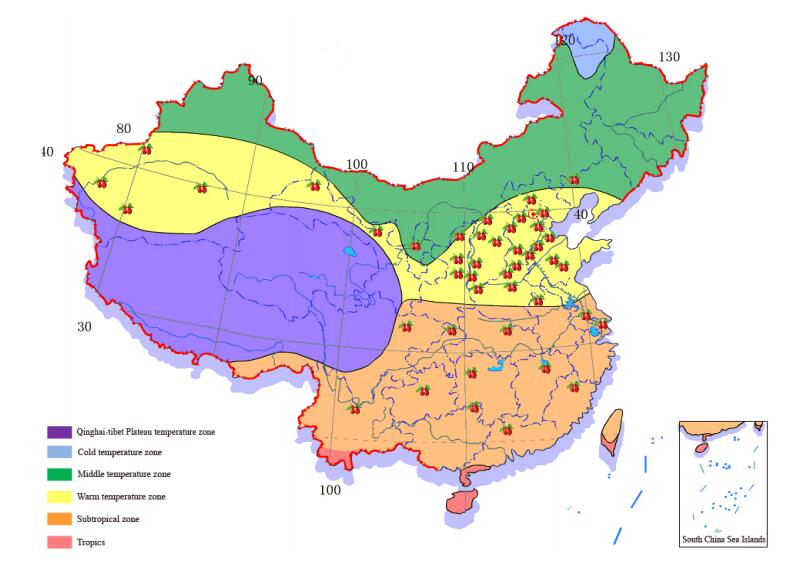 Figure 1. The cultivation regions of Chinese jujube in China.
Figure 1. The cultivation regions of Chinese jujube in China.
Jujube has been introduced into about 50 countries throughout Asia, Europe, Africa, the Americas, and Oceania [9]. However, the global production is heavily concentrated in China, which currently produces more than 99% of the world’s jujubes. In China, the fruit is mostly consumed in the Chinese domestic market. Approximately 3.0% of the Chinese production is exported to the international market, of which 80% is to East and South East Asian countries [6]. Most the exported jujube is dry fruit, followed by candied fruit. Nonetheless, fresh jujube is becoming more popular in both domestic and international markets. South Korea is the second largest jujube-producing country in the world, with a cultivation area of about 5000 ha and annual production around 20,000 tons [9]. Jujube is considered to have good potential in many other countries as a new fruit species, such as in Australia, where its counter-seasonal production to the northern hemisphere provides an opportunity to markets in East Asia [10]. In the United states, jujube was reported as having good potential in regions such as the Southwest, where jujube’s late-spring bud break makes it a good alternative crop to fruits that suffer from late-spring frosts [11,12].
Despite the outstanding adaptability of the jujube tree, cultivation of jujube has many biotic and abiotic production constraints, including diseases, insect pests, cold temperatures and drought [9]. Among the biotic production constraints, diseases caused by fungi and phytoplasmas cause major problems, with an annual economic loss of $4–6 billions [9]. A total of 38 diseases/disease complexes have been reported to infect jujube during pre-and post-harvest stages [13]. There are four major diseases/disease complexes that either occur nationwide in China and/or cause the most yield reduction. These include witches’ broom (associated with phytoplasma presence), leaf-rust (Phakopsora zizyphi-vulgaris), fruit shrink disease (Coniothyrium olivaceum, Alternaria alternata, Dothiorella gregaria, Macrophoma kawatsukai, Phoma destructiva, Fusicoccum sp., and Erwinia jujubovora) and various fruit rot diseases [9,13,14]. Yield reduction caused by these diseases ranges from 10–30% in average years and yield losses can reach 100% in extreme cases. Moreover, some of these diseases, such as fruit shrinkage and fruit rot, also cause quality deterioration in pre-and post-harvest stages, rendering fruits non-edible [14].
Host resistance is expected to be the most efficient way to cope with the above mentioned four major diseases/disease complexes in jujube. A substantial amount of research has been carried out in breeding disease resistant jujube cultivars in China [9,13], and research investment is increasing. Breeding efforts have primarily focused on witch’s broom, fruit shrinkage and various fruit rot diseases. The breeding approaches range from germplasm evaluation, selection of farmer cultivars and introgression of host resistance from wild relatives [7,13]. Nonetheless, an updated analytical summary of these efforts is not available. Moreover, most of this research has been published in Chinese or as informal publications, and thus are not readily accessible by the international community. Herein we summarize the on-going research activities in jujube disease resistance breeding in China, with the aim to identify knowledge gaps that when filled will improve the effectiveness of jujube breeding for enhanced disease resistance. Due to the space limit, we will only review the four major diseases as mentioned above.
Jujube Witches’ BroomJujube witches’ broom (JWB), associated with phytoplasma presence, is an extremely destructive disease commonly found in both North and South China [15], as well as in India [16], Korea [17,18] and Japan [19]. This important and damaging disease has caused yield reductions ranging from 10% to 80% [20,21]. A severely infected tree will die within two to five years [21,22], resulting in million tree deaths by JWB each year in China [22]. Phytoplasmas, or mycoplasma-like organisms (MLO) are plant pathogenic bacteria which lack cell walls and are transmitted by insect vectors. They inhabit phloem; spreading through the plant by way of this vascular tissue [23]. A tree infected with phytoplasmas typically displays severe rosetting, little green leaves and a witches’ broom-like appearance (Figure 2). Based on analysis of ribosomal (16Sr) sequence, Jung et al.(2003) proposed that the phytoplasmas associated with jujube JWB disease in Korea and Japan were designated as a novel, distinct taxon, namely “Candidatus Phytoplasma ziziphi” [18]. The same species was also identified in JWB-infected wild jujube in China [24,25]. The insect vector transmitting JWB is the leafhopper Hishimonus sellatus (Uhler) [21,26,27]. JWB phytoplasmas have been identified solely from jujube trees, which suggested that the feeding preferences of H. sellatus, rather than the resistance of other potential plant hosts may be responsible for this specificity [18].
 Figure 2. A Chinese jujube tree infected by witches’ broom disease. (A) A full branch damaged by witches’ broom disease; (B) A close view of the witches’ broom-like appearance on young shoots.
Figure 2. A Chinese jujube tree infected by witches’ broom disease. (A) A full branch damaged by witches’ broom disease; (B) A close view of the witches’ broom-like appearance on young shoots.
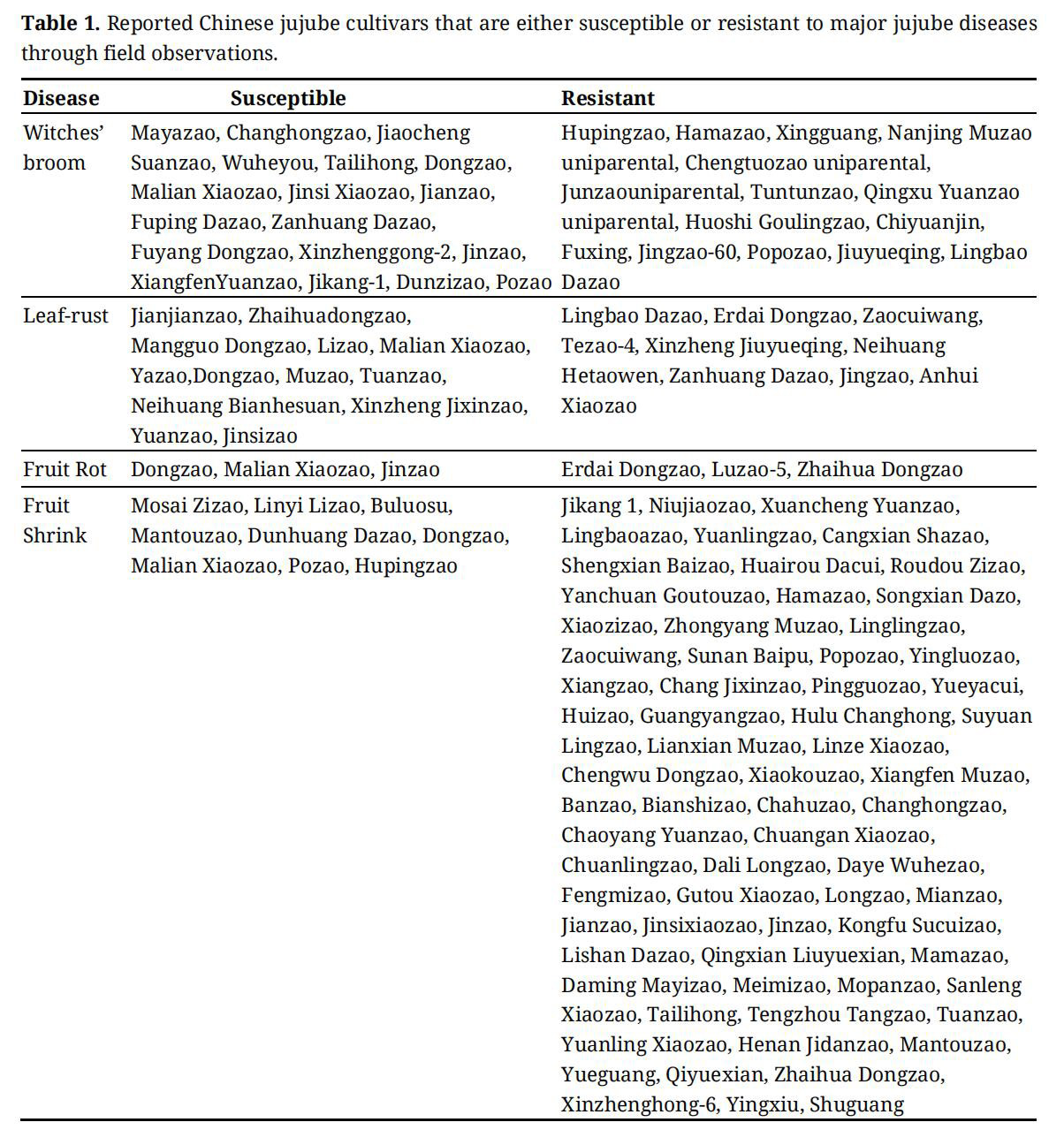 Table 1. Reported Chinese jujube cultivars that are either susceptible or resistant to major jujube diseases through field observations.
Table 1. Reported Chinese jujube cultivars that are either susceptible or resistant to major jujube diseases through field observations.
JWB is the only jujube disease for which breeding efforts have been systematically employed and significant progress has been made [15,28]. Most of the reported breeding activities were carried out in Hebei, China, where JWB has been prevalent [15,28,29]. Resistance was first evaluated in jujube germplasm collections using field resistance as the primary evaluation criteria. Screening for resistance was carried out using grafting methods, where budwood (cutting) was grafted to severely infected adult trees (stocks). A five-grade rating system was applied in the experiment. A large variation of JWB resistance was found in the jujube germplasm evaluated, ranging from highly susceptible to highly resistant [15]. After five years of repeated observations, four resistant clones (T15, J2, J5 and J8) were selected. The selected resistant clones were also evaluated for other agronomic traits such as yield, earliness and postharvest quality. Cultivars with JWB resistance and desirable agronomic traits, such as “Junzao uniparental” and “Hama Zao” have been released and used in production [26]. It is noticeable that in most of these reported studies, the phytoplasma presence was routinely verified using fluorescence microscopy and polymerase chain reaction (PCR) techniques amplifying/sequencing phytoplasma ribosomal sequence (16Sr) using general and specific primers pairs [30,31]. It was reported that resistance to JWB was mainly due to the inhibition of phytoplasma multiplication after infection [15]. Jujube cultivars identified in China as resistant to JWB are presented in Table 1.
Jujube Leaf-RustJujube leaf rust disease, caused by Phakopsora zizyphi-vulgaris (P. Henn), is another serious threat to jujube production [32–34]. Rust mainly occurs on leaves and less commonly on fruits. Since an outbreak of rust disease in Henan, China in 2004, rust has become the most important disease affecting winter jujube production in this region [9]. Based on a survey between 2004 and 2008 in Shandong, China, Yan and Wang reported that rust disease can reduce jujube yield by 10% to 50% [35]. In the 1970, an 85% yield reduction was recorded in Neihuang district, Henan Province, China, during a rust disease outbreak. Since rust disease coincides with fruit development, it also contributes to fruit shrinkage and quality deterioration, and the produced toxic substances can make the fruit inedible [6,35].
It is not clear whether the jujube rust pathogen P. ziziyphi-vulgaris has alternative hosts. Xu et al. found that there was no heteroecism in P. ziziyphi-vulgaris, and summer spores on old and fallen jujube leaves were the primary inoculum source of new infections [33]. Release of spores occurs during rainy periods. The fungus sporulates through the stomata rather than breaking through the epidermis. After spore germination, the pathogen enters the host through leaf stomata. Young leaves are more vulnerable than mature leaves. Small green spots first appear on the blade of the leaves, usually within a week or two after bloom [33,34]. The spots enlarge to one quarter inch or more and yellow droplets (summer spores of the pathogen) appear (Figure 3). Infected trees begin to defoliate in August and September. High moisture and rainfall favor rust disease [14,36].
 Figure 3. Leaf-rust disease (Phakopsora zizyphi-vulgaris) infected leaf of Chinese jujube tree.
Figure 3. Leaf-rust disease (Phakopsora zizyphi-vulgaris) infected leaf of Chinese jujube tree.
Studies on breeding for rust resistance in Chinese jujube are scarce. Li et al. (2010) compared rust resistance in three jujube cultivars including “Dongzao”, “Gagazao” and “Lingbao Dazao” using a leaf disc inoculation method [37]. Resistance in the three tested cultivars varied significantly. “Lingbao Dazao” had highest resistance, whereas “Gagazao” was less resistant and “Dongzao” was the least resistant. Genotype differences in rust resistance was also reported by Gao [38], who evaluated 29 clone selections of “Muzao” and found three clones were resistant, two were moderately resistant, eight were moderately susceptible and 16 were susceptible.
Jujube Fruit Shrinkage Disease ComplexThe jujube fruit shrinkage disease (JFSD) is common in Northern China and it is also reported in Guangxi, Sichuan and Hunan in Southern China (Figure 4). Yield reduction caused by JFSD can reach 80% as reported in Hebei, China, JFSD also causes deterioration of fruit quality due to bitter pulp and early drop [39].
 Figure 4. Fruits of Chinese jujube showing a typical symptom of fruit shrinkage disease. (A) An infected fruit on tree; (B) different type of fruit shrinkage.
Figure 4. Fruits of Chinese jujube showing a typical symptom of fruit shrinkage disease. (A) An infected fruit on tree; (B) different type of fruit shrinkage.
Several kinds of fungi and bacteria have been reported as the causal agents of the JFSD, which infect jujube singly or jointly, making JFSD a disease complex complicating its study and the development of disease management protocols. Xu et al. (1995) found that the causal pathogens of JFSD included three fungal pathogens (Coniothyrium olivaceum, Alternaria alternata, Dothiorella gregaria) and one unidentified bacteria [40]. From 1992 to 1996, the total isolation rates of the four pathogens were 14.9%, 12.69%, 11.47% and 10.66% respectively at the investigated site in Hebei, China [40]. Zhang et al. (2008) reported that there were eight causal pathogens of JFSD, including six fungal pathogens (Coniothyrium olivaceum, Alternaria alternate, Dothiorella gregaria, Macrophoma kawatsukai, Phoma destructiva and Fusicoccum sp.) and two bacteria pathogens (Erwinia jujubovora and one unidentified bacteria species)[39]. However, the pattern of infection by these pathogens and how they interact in causing disease on jujube is not clearly understood.
Large cultivar differences were also found for resistance to JFSD. Xie et al. (2015) screened 268 jujube cultivars for resistance to JFSD [41]. The germplasm included in this study covered three major production regions in Northern China (Changxian, Hebei, Taiyuan, Shanxi, Taigu, Shaanxi). Their result showed that the infected fruits ranged from 0 to 66.2% and disease index ranged from 0 to 55.6% among cultivars. In addition, a significant effect of G × E (both location and year) was detected, suggesting the host resistance was quantitative in nature. Reactions to fruit shrinkage was classified as highly resistant (disease index <5%), resistant (5–10%), moderately resistant (10–15%), susceptible (15–25%) and highly susceptible (>25%).
A screening method using spore suspension inoculation on detached jujube fruit has been developed [42]. Screening results using the spore suspension inoculation method with three pathogens (Alternaria alternata, Phoma destructiva and Fusicoccum spp.) were found consistent with field resistance measurements. Jujube germplasm accessions with high levels of resistance to JFSD were also reported in Hebei [43]. To date, at least 40 jujube cultivars were reported as having resistance to JFSD (Table 1).
Jujube Fruit Rot DiseaseJujube fruit rot disease (JFRD) is also a major constraint in jujube production in China, but like JFSD, JFRD can be caused by different pathogens, either singly or jointly, again suggesting a disease complex.
The causal pathogens include both fungi and bacteria, causing pre- and post-harvest fruit rots. When infections occur involving single pathogens, associated common disease names are often applied. Commonly reported pathogens include Colletotrichum gloeosporioides (Anthracnose fruit rot), Alternaria spp. (Alternaria fruit rot), Phytophthora spp. (Phytophthora fruit rot), Fusarium spp. (Fusarium fruit rot), Erwinia Jujubovora and Xanthomonas arboricola (Bacterial fruit rot).
Anthracnose fruit rot (Figure 5) has been reported in all jujube production regions in China, except in the provinces of Xinjiang, Ningxia and Liaoning [43]. Anthracnose typically starts when fruit reach the ripening stage. Fallen leaves provide the primary source of infection while conidia act as secondary sources and are disseminated by rain [44]. Yield losses of 20–30% have been commonly reported in infected trees, but in years of severe disease, yield losses can reach 80%, as described in Shanxi, Shannxi, Henan, Shandong et al. in 2012 [45]. Moreover, the pathogen remains inside the fruits causing rot and decay of dry fruits during storage. The damage rate can reach 50% for dry fruits [43].
 Figure 5. Chinese jujube infected by Anthracnose fruit rot disease (Colletotrichum truncatum). (A) An infected fruit; (B) Infected leaves.
Figure 5. Chinese jujube infected by Anthracnose fruit rot disease (Colletotrichum truncatum). (A) An infected fruit; (B) Infected leaves.
Alternaria fruit rot, also known as black spot or black mold rot, caused by Alternaria alternata, is the most important disease causing latent infections and serious postharvest losses of fresh and dry fruits [25,34]. This pathogen also causes leaf spot disease in Xinjiang [46]. Other Alternaria species, including Alternaria yaliinficiens R.G. Roberts and Alternaria tenuissima (Fr.) Wiltshire, have also been reported to cause fruit rot in China [47]. Since Alternaria spp. also causes JFSD, as mentioned above, the relationship between Alternaria fruit rot and fruit shrinkage in jujube needs more detailed investigations. In this pathogen and some other pathogens, there is a possibility that JFSD and JFRD are variations of the same disease but based on different symptoms.
At least three species of Phytophthora including Phytophthora citricola, P. nicotianae and P. palmivora, have been reported to cause pre-harvest fruit rot of jujube in Korea [32,48]. However, so far, these pathogens have not been officially reported in China. Fusarium proliferatum and Fusarium oxysporum have been reported as the pathogens causing jujube soft rot disease in Henan province, China [43,47].
Different fruit rot diseases have been reported in Korea and China. The causal agents include Xanthomonas arboricola, Rhizopus stolonifer, Penicillium expansum and Erwinia jujubovora, of which, the latter was also reported to causes JFSD [39,49].
Despite the importance of fruit rot disease, there have been few reported studies searching for host resistance in jujube. Li et al. (2016) observed large cultivar differences in resistance to black spot caused by Alternaria alternata [50]. The infection rate was above 30% in cultivar “Junzao” and “Hupingzao”, whereas no symptoms were found in cultivar “Mayazao” and “Suanzao”. This result indicates that it is possible to select host resistance to Alternaria fruit rot in jujube and similar studies could be carried out with other fruit rot pathogens.
Knowledge about species, races, and spatial and temporal occurrence of the major diseases remains deficient in major jujube production regions. It is speculated that more species may be present on jujube farms than have been reported [13]. Accurate identification of the pathogens is essential to manage diseases, and this can be achieved through the combined use of morphological or molecular techniques [31,51]. For example, in situations where mixed pathogens may synergistically infect the jujube tree, next generation sequencing can be applied to clarify the pathogens involved. Draft genomes of these major pathogens can be developed using this same approach.
Knowledge and understanding of the species origins, dispersal and genetic diversity should be investigated by the combined use of morphological, biochemical, molecular and pathology approaches. So far, pathogen race designations for these pathogens are lacking but, as sources of resistance are better characterized, development of race panels should become possible. Comprehensive understanding of pathogen genetic variability is critical to germplasm enhancement and development of new cultivars, as well as for deployment of sources of resistance in the field. In the case of diseases with race specificity, such as leaf rust, this is especially useful. Furthermore, it is important to identify alternative hosts of these major jujube diseases. The impact of alternative hosts on disease development of jujube trees has rarely been studied. The majority of the pathogen species causing disease on jujube are not host-specific. Therefore, other hosts that are present within a region could harbor and perpetuate pathogens during unfavorable conditions in jujube fields.
Despite the identification of various resistant genotypes in jujube germplasm (Table 1), there is still a lack of consensus concerning the repeatability and durability of disease resistance. This results from the evaluation of resistance being based mainly on field observation of infected trees, which offers only an approximate indication of the true susceptibility, and that is limited by the pathogen’s genetic diversity in the locations studied. Crop infection in fields, especially in perennial crops, is heterogeneously distributed and is influenced in part by genotype, microclimate, inoculum and cropping system. For example, it has been observed that seasonal variation strongly influenced the distribution of jujube rust disease incidence in fields at Hebei, suggesting that the observational results may need to be confirmed with the aid of artificial inoculation [33]. Many techniques have been developed and used to assess the disease susceptibility of the different tissues of other major fruits in the laboratory, as well as in the field, and on detached and/or attached fruits.
Due to the lack of consistent inocula, screening results continue to be inconsistent across laboratories and locations. The inoculum type and mode of application has differed depending on the researchers at different laboratories and regions. It is imperative, therefore, to develop reliable and consistent screening methodologies that will clearly identify the various levels of resistances that may exist in germplasm collections.
Furthermore, standardization of factors of host, pathogen, and environment that affect infection success is critical to the development of good screening methods that are unambiguous in differentiating among the various levels of resistance in different tissues and organs. Variation in such factors (host, pathogen, and environment) can affect the sensitivity and reproducibility of the screening method and hence need to be optimized. Tissue (including attached vs detached), age, field vs lab evaluation, and their roles in screening results must be standardized. Establishing standard details of screening methods across research programs would greatly increase the usefulness of results to the jujube research community.
In addition, to standardizing the screening of disease resistance, it is paramount to develop diagnostic tools to detect disease infection. At present, diagnostic tools that are available are not capable of detecting all the pathogen strains present across different regions. This situation has contributed to the slow progress in the development of resistant germplasm because breeders do not have accurate information about the causal agents of these complex diseases, such as JFSD and JFRD.
Despite the large number of available germplasm accessions, jujube has a few improved cultivars in comparison to other major fruit trees. So far most of the cultivars used in production are landraces and traditional cultivars historically selected by farmers [6,52]. As summarized above, jujube breeding is still in a primitive stage and host resistance against various species and isolates of pathogens have not been clearly identified, especially in the cases of JFSD and JFRD. In addition, there are several major knowledge gaps in jujube genetics and breeding method, which complicate the development of disease resistant jujube cultivars.
Cultivar MislabelingThere are more than 900 jujube cultivars in China, which vary widely in traits such as leaf shape, fruit form, size, color, flavor, keeping quality and fruiting season [6]. These cultivars have been traditionally classified based on morphological characteristics [5,52]. Jujube has a long history of being introduced and exchanged among different regions in China and different countries and continents. However, records and labels of the cultivars have not always followed the same naming conventions, and many accessions have limited information about their correct identity. Therefore, homonyms and synonyms are common among the names of jujube cultivars, and this restricts the sharing of correct information and materials among jujube researchers and hampers the use of jujube germplasm in breeding. Xu et al. demonstrated that there was a high level of synonymous mislabeling (duplicates) in Chinese jujube germplasm, accounting for about half of the collection (47.3% of the 947 accessions) [53]. Song [54] used SNP markers to assess the genetic identity of 114 jujube cultivars and detected 17 synonymous groups, including 49 cultivars. Misidentified parentage was also identified in the same experiment.
Therefore, the lack of accurate information on jujube genetic identity and pedigree relationship hinders the exchange of information among breeders. Misidentification of cultivars can be minimized by DNA fingerprinting technology [53]. Consistency of resistance evaluations can be improved by using only plants that have been previously genotyped with molecular markers.
Understanding about the Reproductive Biology and Mating SystemsThe current breeding scheme of Chinese jujube largely relies on clone selection from natural progenies of landraces and very few from controlled crosses [25]. One of the barriers in breeding is that jujube has very small flowers (approximately 5–7 mm in diameter) making emasculation difficult. Jujube also has a very low natural fruit set (1%) and suffers a high percentage of embryo abortion. The probability of obtaining hybrids is only around 0.01% using conventional methods [7]. This failure is largely due to the lack of comprehensive understanding about the reproductive biology and mating system in Chinese jujube germplasm, especially as regarding to self-incompatibility and male sterility.
Asatryan and Tel-Zur (2013) analyzed the self-incompatibility system in three Ziziphus species and tested whether autonomous self-pollination, parthenocarpy or agamospermy occurs [4]. They reported that the self-incompatibility system operating in the studied Ziziphus species is gametophytically controlled. Yao et al. (2013) examined the blooming types [13], flowering characteristics, self-pollination/self-fertility, and seed development of 56 jujube cultivars in New Mexico, United States. They reported that out of the 56 tested cultivars, seven were self-pollinating/self-fruitful in New Mexico. Most cultivars had better fruit set from open pollination than self-pollination with a few exceptions (e.g., “Li”, “Li-2”, and “Redland”). The report of Yao et al. (2013) differed from Liu and Wang (2009) in which a high level (87.8%) of self-fruitful cultivars was observed, based on an experiment including 179 cultivars [1,13]. Here is a clear example where a comprehensive understanding and validation of the germplasm used in two studies would greatly improve the interpretation of their combined results.
Recently, with the assistance of genomic approaches, new achievements have been made in understanding the molecular mechanism of self-compatibility in jujube. Based on the result of re-sequencing 31 accessions of cultivated and wild jujubes (Ziziphus jujuba var. spinosa), the genetic mechanism of GSI in jujube was demonstrated to be the same as that found in several other Ziziphus species [4,8]. Parents sharing the same S haplotype often result in seedless jujube kernels [8].
Candidate S-RNase gene (S1) and S-like RNases that belong to the T2-RNase family were identified in the genomes of “Junzao” and “Dongzao” [8]. A transcriptome analysis revealed that the S1 gene was specifically expressed in flowers, and the S-like RNase genes were expressed in all tested tissues. Based on 21 SNPs and 3 indels, Huang et al. (2016) found that S1 was the most common haplotype in Chinese jujube [8]. They suggested that by crossing two different S haplotypes (i.e., “Dongzao” × “Linyi Lizao” and “Dongzao” × “Zhongning Yuanzao”), the incompatibility could be overcome. This progress shed light on marker-assisted germplasm evaluation for self-compatibility in jujube. However, this new knowledge has not been practically applied in most jujube breeding programs. New breeding methods, such as marker-assisted germplasm screening for incompatibility will likely have significant impact in jujube breeding and cultivar deployment in the future.
Parallel to the progress in understanding self-incompatibility, a breakthrough has also been made in overcoming the barrier of cross-hybridization using male sterility [6]. Male sterility has been found in at least five Chinese jujube cultivars so far, which provides a key solution for the difficulty in flower emasculation and embryo abortion. In male sterile cultivars, microspores massed and became disaggregated after tetrads so the pollen became abortive. By employing male sterility in controlled crosses, Liu et al. [28] developed a method that combined selection of parental cultivars and planting or grafting parents together with a net covering during the flowering season (Figure 6). Honey bees were used to carry out the controlled pollination. Culture or rescue of young embryos, and molecular identification of hybrids followed as necessary. This strategy was used to obtain hybrid jujube populations [55,56].
 Figure 6. Production of hybrid progenies using controlled crosses in Chinese jujube breeding. (A) Bi-parental controlled cross using net bags; (B) Bi-parental controlled cross using male-sterile parental lines.
Figure 6. Production of hybrid progenies using controlled crosses in Chinese jujube breeding. (A) Bi-parental controlled cross using net bags; (B) Bi-parental controlled cross using male-sterile parental lines.
Selection of parent combinations is the key for emasculation‐free cross breeding in Chinese jujube. Based on evaluation of the reproductive traits of approximate 500 genotypes of Chinese jujube, Liu et al. (2006) screened out several genotypes suitable for use as emasculation-free female parents that are male sterile but outcross fertile [9], self‐unfruitful but outcross fertile, or self‐sterile but outcross fertile [35,57], in addition to some excellent genotypes suitable for use as male parents with rich pollen of high viability [9,57]. Nevertheless, in breeding for disease resistance, there remains a lengthy process to combine diseases resistance with male sterility to obtain hybrid progenies with disease resistance. More male sterile candidate parents need to be identified to broaden the genetic background in breeding programs and marker-assisted selection should be practiced to shorten the selection progress.
Progress in Development of Genomic Tools for Marker-Assisted BreedingThe importance of Chinese jujube warrants greatly expanded efforts in developing genomic tools for supporting germplasm management and breeding. However, progress in developing genomic tools for jujube is limited in general when compared to that achieved for temperate fruit trees and annual crops. Some noticeable progress has been made over the last five years, including the development of jujube draft genomes and constructions of linkage maps. With the sequencing of jujube cultivars “Dongzao” and “Junzao” [8,28], an immense number of molecular tools have become available. Thousands of SNP markers have been identified and are currently being used by breeders in the development of new cultivars.
The draft genome of a cultivar for fresh consumption of Chinese jujube, “Dongzao”, was developed using next-generation-sequencing (NGS) and a novel combination of BAC-to-BAC (or hierarchical shotgun sequencing) and mate-paired libraries of various sizes from whole-genome-shotgun sequencing (WGS)[28]. The final assembly spanned 437.65 Mb (98.6% of the estimated genome sequence) with 321.45 Mb anchored to the 12 pseudo-chromosomes. A large proportion of the “Dongzao” assembly (73.6%) was anchored to a genetic map of 2419 SNP markers obtained using restriction-site-associated DNA sequencing (RAD-seq) that covered all 12 chromosomes. In total, 32,808 gene models were predicted from the “Dongzao” genome assembly [28].
A draft genome of a “dry” jujube cultivar, “Junzao”, was developed by Huang et al. (2016)[8], together with resequencing the genome of 31 accessions of geographically-diverse cultivated and wild jujubes (Ziziphus jujuba var. spinosa). Compared to the genome of the jujube cultivar “Junzao”, the genome of the cultivar “Dongzao” was ~86.5 Mb larger. In part this was due to transposable elements inserted in the “Dongzao” genome.
So far only two high density linkage maps in Chinese jujube have been published. Zhang et al. (2016) constructed a map from an inter-specific F1 population [29], using restriction-site associated DNA (RAD) markers. The mapping population, including 105 F1 progenies, was generated between parents Z. jujuba “JMS2” × Z. jujuba var. spinosa “Xing 16”. Out of the 2872 RAD markers used for map construction, 1307 were linked to the female (Z. jujuba) genetic map, 1336 to the male (Z. acidojujuba) map, and 2748 to the integrated map, which spanned 913.87 centimorgans (cM) and had a 0.34 cM average marker interval. There were 12 linkage groups (LGs) in the integrated map. Using the genotyping-by-sequencing (GBS) strategy, Zhang et al. (2016)[42] reported a high-density genetic map based on a F1 population with 145 progenies and their parents (“Dongzao” × “Zhongning Yuanzao”). The map was based 2540 SNP markers distributed among 12 linkage groups, and spanned 1456.5 cM. The average marker interval was 0.88 cM. The genetic map enabled the anchoring 224Mb (63.7%) of scaffolds from the sequenced “Junzao” genome.
Despite the rapid progress in genomic research, to date, no QTLs associated with diseases resistance have been discovered across the jujube genome. In addition, the development of methodologies such as genome-wide association studies and genomics selection could be used in the screening of accessions available in germplasm collections, as well as in farmer’s fields, to discover new sources of genes for resistance against diseases in jujube, as well as genes associated with good horticultural and quality traits. These novel genes could then be incorporated into ongoing breeding programs.
Several knowledge gaps remain to be filled in the current breeding scheme for disease resistance. Firstly, there is a lack of understanding about the major jujube diseases, regarding the associated pathogen’s taxonomy, genetic diversity and geographical distribution. Secondly, there is a lack of effective and robust methods for screening diseases resistance. A standardized phenotyping protocol needs to be established and adopted to ensure consistent screening results comparable across locations and differing breeding programs. Thirdly, in the aspect of breeding method, the mislabeling rate is high in the jujube germplasm due to the mistakes during the long history of cultivar introduction and planting material propagation. These errors greatly reduce the accuracy and efficiency of breeding programs, restricting progress in germplasm enhancement.
Although improvements have been made, in general there remains an insufficient understanding of the reproductive biology and mating system in Chinese jujube. A bottle neck in jujube breeding is the difficulty in making crosses and producing hybrid progenies. Despite the many jujube breeding programs, the total number of crosses (both open pollination and bi-parental crosses) and the size of evaluated progenies (or families) has been small. The reproductive biology, specifically cross- and self-incompatibility and male sterility of the jujube germplasm involved, has not been systematically investigated for most breeding programs. Therefore, controlled crosses often have low fertilization rate and aborted seeds.
Last but not the least, our understanding of the genetic basis for host resistance in jujube is limited. Detailed information of the different aspects of resistance genes and QTLs, their original sources, their distribution in the key parental cultivars, as well as in the primary gene pool of Chinese jujube is yet to be generated and documented. This work needs to be carried out by research teams with expertise in both plant breeding and plant pathology. Unfortunately, a lack of participation from plant pathologists is a common weakness in most jujube breeding programs.
Recent progress in the discovery and utilization of male sterility should significantly enhance the capacity to produce hybrid progenies. If properly exploited, this should speed up the introgression of disease resistance into parental lines. Moreover, the rapid progress in developing jujube genomic tools should enable marker-assisted breeding, which can accelerate the enhancement and introgression of host resistances. These two major breakthroughs have the potential to put jujube breeding on a new platform. However, the above-mentioned knowledge gaps, especially the lack of understanding concerning the pathogens involved and host resistance in general, need to be filled before these advanced genomic tools can be practically applied to assist jujube breeding.
The authors declare that they have no conflicts of interest.
This research was partially supported by the National Natural Science Foundation of China (Grant No. 31760203) and the Key Innovation Projects of First-Class University in Western China (Grant No. ZKZD2017008).
The authors would like to thank Dr. Sue Mischke for reviewing and editing of this manuscript, Professors Zhanwen Li and Baiyun Li for providing pictures of jujube diseases.
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
41.
42.
43.
44.
45.
46.
47.
48.
49.
50.
51.
52.
53.
54.
55.
56.
57.
Song L, Meinhardt LW, Bailey B, Zhang D. Genetic Improvement of Chinese Jujube for Disease Resistances: Status, Knowledge Gaps and Research Needs. Crop Breed Genet Genom. 2019;1:e190015. https://doi.org/10.20900/cbgg20190015

Copyright © 2020 Hapres Co., Ltd. Privacy Policy | Terms and Conditions




