Location: Home >> Detail
Immunometabolism. 2019;1:e190009. https://doi.org/10.20900/immunometab20190009

Center for Immunology, Department of Laboratory Medicine and Pathology, University of Minnesota, Minneapolis, Minnesota 55455, USA
* Correspondence: Henrique Borges da Silva.
This article belongs to the Virtual Special Issue "T Cell Metabolic Reprogramming"
Following activation, CD8 T cells transition from reliance on mitochondrial respiration to increasing utilization of aerobic glycolysis. After the effector phase, however, reversion to mitochondrial metabolism is pivotal generating memory CD8 T cells. We recently showed that sensing of extracellular ATP (eATP) through the receptor P2RX7 is crucial for both production and the long-term survival of memory CD8 T cells, evidently through promoting mitochondrial maintenance. Unexpectedly, these results indicated that sustained P2RX7 activation is required for memory CD8 T cell homeostasis, suggesting constant exposure to eATP, in contrast with the proposed role of eATP as an acute “danger” signal released by dying cells. Active release through transmembrane channels is another path for eATP export. Indeed, CD8 T cells express Pannexin 1 (Panx1) which has a reported eATP release function in vitro and is itself induced by P2RX7 and/or TCR engagement. Such a role for Panx1 could potentially provide a feed-forward mechanism for cell-autonomous P2RX7 signaling. This model envisages that memory CD8 T cells maintain themselves at the cost of reduced intracellular ATP levels, which at first glance would seem to be detrimental for sustained T cell maintenance. On the other hand, the need to tightly regulate levels of intracellular ATP may be critical for the durability and adaptability of memory CD8 T cells, hence engagement of the P2RX7/Panx1 axis may allow these cells to fine tune their metabolic status to meet changing demands. In this Perspective, we discuss how this pathway may influence memory T cell maintenance.
Cytotoxic CD8 T cells are crucial to control viral infections and tumors [1,2], and promoting durable memory CD8 T cells is a goal of immunizations. During the course of antigen-driven immune responses, CD8 T cells undergo diverse changes in their biology at transcriptional, epigenetic and functional levels [1,3–8]. Together with such components, cellular metabolism is dynamically re-wired throughout the CD8 T cell response. At steady-state, naïve CD8 T cells are metabolically quiescent, primarily using oxidative phosphorylation (OXPHOS) [8] and circulate through lymphoid organs receiving homeostatic signals. This occurs until cognate antigen presentation by antigen-presenting cells (APCs) engages the T cell receptor (TCR) which, together with costimulatory signals, induces a metabolic shift towards preferential anabolic growth and biomass production, highlighted by increased engagement of aerobic glycolysis [8–10]. Consequently, effector antigen-specific CD8 T cells undergo rapid and extensive proliferation. At the end of this “effector” phase, most CD8 T cells succumb to apoptotic death but a small population of memory precursors emerges [11]. Memory CD8 T cells undergo a subsequent remodeling of their metabolism, having increased respiratory levels and spare respiratory capacity compared to naïve or effector counterparts [12–14]. This metabolic re-wiring is fundamental for long-term survival, self-renewal and for the potential of memory CD8 T cells to quickly respond to antigen re-exposure [1].
Extensive research has been done in the past years to understand the basis for metabolic programming of memory CD8 T cells. The acquisition of a memory transcriptional program is evidently important. Transcription factors can directly promote memory precursor differentiation [15], leading to increased expression of mitochondrial-related genes [13,16–18]. Conversely, metabolic alterations can promote memory CD8 T cell differentiation [8], such as negative modulation of the mechanistic target of rapamycin (mTOR) signaling pathway [19] and activation of the adenosine monophosphate-activated protein kinase (AMPK) pathway [20]. These metabolic alterations depend on cell-intrinsic features, such as asymmetric division during effector phase [21,22], and in part on cell-extrinsic signals. Among extracellular signals, metabolites released by autocrine or paracrine cell sources are ubiquitously present and can play a very important role. We have recently found that sensing of one of these metabolites, extracellular adenosine triphosphate (eATP), by the purinergic receptor P2RX7 is required for memory CD8 T cells to establish and to acquire optimal mitochondrial metabolism [23]. Whether eATP primarily acts through a cell-extrinsic or cell-intrinsic pathway in its action on memory CD8 T cells is unclear and is discussed in this Perspective. P2RX7 is differentially expressed by distinct memory CD8 T cell subsets. Memory CD8 T cells can be divided into recirculating and non-recirculating subsets. Among circulating memory CD8 T cells, Central memory T cells (TCM) traffic through the blood, lymph and secondary lymphoid organs (SLOs), canonical effector memory T cells (TEM) can enter non-lymphoid and some lymphoid tissues from the blood and drain via the lymphatics. Conversely, “tissue-resident” memory T cells (TRM) do not recirculate at all, instead being maintained long-term in lymphoid and non-lymphoid sites [24,25]. TCM express higher P2RX7 than TEM, but TRM express the highest levels of this receptor [23,26,27]. These differences in P2RX7 expression level may have important biological consequences for its function in distinct memory T cell subsets. In this article, we primarily focus on circulating memory CD8 T cells, but briefly discuss implications for tissue-resident memory CD8 T cells.
Most of the findings reported above, including our own, focused on the understanding of factors that promote generation of memory CD8 T cells; however, the signals that are necessary for memory CD8 T cells to maintain their feature characteristics and to survive long-term are less well understood. There is some evidence, for instance, that both the phenotype and survival of memory CD8 T cells is constantly reinforced. Among cell-intrinsic factors, sustained function of Foxo1 maintains long-lived memory CD8 T cells [28], while constant Id2 keeps the effector memory signature phenotype [29]. Extracellular signals, such as the cytokines Interleukin (IL)-7 and IL-15 also play a key role [30–32]. Notably, we observed that sustained P2RX7 signaling is required for long-term survival of memory CD8 T cells [23]. In this Perspective, we will focus on the source and role of sustained eATP as an extracellular signal to maintain the function and survival of memory CD8 T cell function and survival. In particular, we discuss the potential role of autocrine eATP as a dominant source of the P2RX7 stimulus once infection has been controlled and inflammation receded. We also discuss the consequences of an ATP export-eATP recognition axis for regulating metabolic pathways in memory CD8 T cells, and propose a hypothetic model predicting how eATP sensing influences the generation and the long-term survival of memory CD8 T cells.
ATP is the quintessential cell fuel, driving virtually all cell functions that require energy. Typically, intracellular ATP concentrations vary between 3 and 10 mM [33]. Conversely, the concentration of eATP at steady-state is very low, at the nanomolar range [34]. However, in response to infections or insults, passive or active release of ATP into the extracellular microenvironment increases eATP concentrations into the hundred micromolar range [35]. In mammals, extracellular nucleotides are recognized by purinergic receptors. These receptors can be roughly divided into adenosine-recognizing, G-protein coupled P1 receptors and P2 receptors, which bind to eATP, eADP or similar nucleotides [36,37]. Receptors for eATP include the P2RX family, which are gated ion channels. All of the P2RX receptors can induce cation exchange (including Ca2+ influx) following eATP engagement, but P2RX7 has unique characteristics [38] and will be the focus of this Perspective. First, P2RX7 uniquely carries a long C-terminal region which interacts with several intracellular proteins [39]; second, P2RX7 has low affinity for eATP, its activation only occurring in the presence of ~100 μM eATP [38]; third, sustained or intense activation of P2RX7 by high concentrations of eATP (or through ADP-ribosylation by the nicotinamide adenine dinucleotide (NAD+)-activated enzyme ARTC2.2, in murine cells) can lead to non-specific pore formation and subsequent cell death [38,40]. In T cells, together with TCR stimulation and costimulation, eATP sensing play important roles in initial responses such as CD27 or CD62L shedding [41,42], IL-2 and IFN-γ production [43], or cell proliferation [44–46]. Importantly, while multiple P2RX receptors are expressed in mouse and human lymphocytes, many of the functional consequences of eATP signaling in these cells can be attributed to P2RX7, specifically [38]. Mechanistically, P2RX7 stimulation engages an array of intracellular signaling and metabolic pathways, which control proliferation and activation in various cell types—these are well-described in recent reviews [38,47,48], and are not further discussed here in the interests of space. However, eATP stimulation of P2RX7 can also promote cell death of lymphocytes via formation of the non-specific pore discussed earlier [38,49]. Indeed, starting with early studies, the same investigators have often reported both stimulation of lymphocyte proliferation and stimulation of lymphocyte cell death by eATP and P2RX7 [44,45,50,51]. This partly reflects lymphocyte heterogeneity, but also the difficulty of modeling physiological eATP encounter through in vitro assays, making it hazardous to rely on such studies to try to predict the impact of P2RX7 stimulation on lymphocyte populations in vivo.
Indeed, during in vivo immune responses, P2RX7 has been found to play either a negative or a positive role in cell function and homeostasis depending on the T cell subset studied. For example, in CD4 follicular helper and regulatory T cell populations, P2RX7 stimulation predominately leads to pore-induced cell death [52–54]. Conversely, P2RX7 can promote generation of T-helper 1 (Th1) [54] and Th17 cells [55,56]. Most of these studies did not track antigen-specific CD4 T cell responses, and future research focusing on antigen-specific CD4 T cells will be necessary to better understand how eATP sensing synergizes with cognate antigen recognition. In contrast, less was known about the in vivo role of P2RX7 in CD8 T cells. Recent works from our group and others show high expression of this receptor by tissue-resident memory CD8 renders them susceptible to cell death induced by sterile damage and subsequent release of high eATP and especially NAD [26,27,57]. More importantly, however, we demonstrated that, in response to acute viral infections, eATP sensing through P2RX7 promotes the generation, homeostasis and function of circulating and resident memory CD8 T cells [23]. Mechanistically, we found that P2RX7 deficiency causes loss of mitochondrial mass and function in memory precursor and long-lived memory CD8 T cells. This aligns with previous studies showing that transfection of cell lines with P2RX7 increased their mitochondrial health [58], and that P2RX7 expression is required for mitochondrial maintenance in microglia [59]; at the same time, these are other reports found that strong stimulation of P2RX7 led to mitochondrial Ca2+ overload, ultimately resulting in fragmentation of the mitochondrial network and cell death [58,60]. Once again, such findings highlight the notion that P2RX7 is a two-edged sword, promoting mitochondrial health and cell growth or causing mitochondrial damage and cell death, depending on the intensity of the eATP encounter. Our studies indicate that, in vivo, P2RX7 is beneficial to activated CD8+ T cells and supports their mitochondrial maintenance [23] suggesting that this is the dominant physiological role, at least for this T cell subset.
Our results have also uncovered an interesting aspect of this pathway, in which sensing of eATP evidently needs to be sustained for the long-term survival of memory CD8 T cells [23] (unpublished data). In the context of acute antigen exposure, eATP can be released by inflamed and/or dying cells from the surrounding microenvironment [38], making it plausible effector and memory precursor CD8 T cells are in the presence of sufficient eATP for P2RX7 engagement. After acute infection is resolved and inflammation recedes, however, it might be expected that sources of paracrine eATP would diminish. If P2RX7 activation is constantly required for long-lived memory CD8 T cells, then an important question arises: what are the eATP sources after the inflammation-promoting antigen is cleared?
As mentioned before, ATP can be released into the extracellular microenvironment roughly by two distinct paths: either “passive” release by inflamed and/or dying cells, or “active” release via cell membrane transporters [61]. During an ongoing viral infection, infected cells will likely release ATP in a non-specific, passive way, especially cells that undergo necrotic cell death [62]. Apoptotic cells can also release ATP; however, they can use cell membrane transporters to do so. One particular transporter is used by apoptotic cells: Pannexin 1 (Panx1) [63]. Panx1 is part of the pannexin hemichannel family and is broadly expressed by immune and epithelial cells [64]. In apoptotic cells, Caspase 3 mediates the activation and opening of Panx1 [65] which is understood as part of the normal apoptosis process [66]. Additionally, Panx1 can be activated transiently, in a caspase-independent process, in healthy cells [67]. This can be induced by several signals such as chemokine binding to CCR5 or CXCR4 [68], adrenoreceptor binding [67], mechanical stresses [65] or cell membrane depolarization [69].
Notably, Panx1 is also expressed by T cell subsets, including human Jurkat T cells [63], murine thymocytes [66] and mature CD4 and CD8 T cells [70]. This suggests CD4 and CD8 T cells may have the intrinsic ability to release ATP, in a Panx1-mediated way. In the past decade, several studies have tested this hypothesis using in vitro systems. In these systems, after TCR stimulation and Ca2+-mediated ATP production by mitochondria, Panx1 quickly mediates a first wave of ATP release, which can activate P2RX7 (and other eATP receptors) in an autocrine feedback loop [71,72]. This autocrine P2RX7 stimulation can enhance TCR stimulation, IL-2 production and T cell proliferation [46,71,73]. Indeed, it has been shown in vitro that Panx1 increases pericellular eATP concentrations in the micromolar range, while concentrations further from the cell membrane are considerably lower. This leads to a local eATP “cloud” that could ensure autocrine P2RX7 activation in T cells, even in the absence of extrinsic sources of eATP [46,73]. It is therefore not unreasonable to imagine similar processes happen in an in vivo response to antigen, although no direct evidence of this currently exists. Through such a mechanism, CD8 T cells would have the potential to create their own eATP pool, at least in part via Panx1 function. In contrast with acute infection—where eATP coming from the inflamed tissue will likely be dominant, this autocrine pathway might be relevant for long-term maintenance of memory CD8 T cells after infection and inflammation have subsided. In our previous report [23], we used IL-15 in vitro polarization of CD8 T cells in the absence of cognate antigen, which induced them to acquire phenotypic and metabolic characteristics of memory CD8 T cells [13,74]. In this system, Panx1 blockade abrogates memory differentiation and mitochondrial metabolism of CD8 T cells, in a P2RX7-dependent way and correlating with decreased eATP [23]. This suggests that in a microenvironment with physiologic low levels of eATP, Panx1 mediates CD8 T cell-autocrine release of eATP which in turn is sensed by P2RX7, and this possibly plays a role in maintaining the phenotype and metabolism of memory CD8 T cells. The role of CD8 T cell-intrinsic Panx1 in the establishment and maintenance of memory CD8 T cells is under investigation, and such studies will shed light on whether this pathway has a biological significance for CD8 T cell responses. The P2RX7 channel itself may also mediate cell-autocrine eATP release, as suggested by reports using P2RX7-transfected cell lines [75] and P2RX7-null tumor bearing mice [76]—although whether P2RX7 directly exports eATP export or rather indirectly enhances activation of Panx1 or other ATP transporters is not clear, and should be a subject of future research.
One important question that arises in light of the potential role of Panx1 for memory CD8 T cells is what advantage would be offered by CD8 T cells exporting ATP for their own sensing, in a seemingly futile cycle. A direct consequence would be ensuring memory CD8 T cells perpetuate a level of Ca2+ influx, driven by P2RX7 stimulation in response to autocrine eATP: maintaining mitochondrial Ca2+ levels is an important factor in promoting mitochondrial health [77], and so this could be a self-preservation mechanism for ensuring the sustained ability of memory T cells to undergo OXPHOS. Alternatively, and more speculatively, the act of exporting ATP itself might have important consequences for memory CD8 T cells maintenance. Memory CD8 T cells typically have increased mitochondrial content (compared to naïve and effector cells), as well as increased baseline and stimulated mitochondrial respiration rates [8,13,78]. Given one of the end products of OXPHOS is ATP production, memory CD8 T cells are expected to produce increased amounts of ATP on a per-cell basis. ATP is used for bioenergetic and biomass production processes, but memory CD8 T cells are metabolically quiescent [8]. Therefore, a potential consequence of sustained OXPHOS could be excessively high levels of intracellular ATP in memory CD8 T cells. While this may be valuable to the cell in terms of an “energy reserve”, there may also be negative consequences. For example, ATP has a high affinity for magnesium ions (Mg2+) [79], hence an aberrant increase in intracellular ATP stocks could lead to a depletion in cytosolic free Mg2+ levels. Mg2+ is an essential cofactor for diverse cellular processes, especially protein translation [80], hence dysregulated levels of intracellular ATP could lead to dire consequences for the overall cellular activity. This is the case for Salmonella bacterial cells where high intracellular ATP concentrations induce cell growth arrest in the presence of low Mg2+ culture conditions [81]. In CD8 T cells, Mg2+ is fundamental for the ability of performing cytotoxic functions during chronic viral infections [82]. Memory CD8 T cells are by definition able to quickly reactivate and perform such cytotoxic functions upon antigen challenge [1]. Thus, Panx1-mediated ATP export may also be important to prevent excessive ATP accumulation and subsequent Mg2+ chelation, which would hinder the ability of memory CD8 T cells to respond to a secondary infection. In this way, one might consider Panx1 activity (sustained by P2RX7 activation) serving as a “release valve” to balance the levels of intracellular ATP.
Panx1-mediated ATP export by memory CD8 T cells may also have consequences for the function of the AMPK-mTOR axis. Studies suggest initial memory CD8 T cell generation is favored by activation of the AMPK signaling pathway [20,83,84] and inhibition of mTOR signaling [19,20,85–87]. AMPK can be activated by a Ca2+ influx-dependent pathway, and Ca2+ influx can be directly controlled by P2RX7. AMPK can also be activated by an LKB-dependent pathway, where a high AMP/ATP ratio promotes its activation and subsequent mitochondrial biogenesis and optimal function in CD8 T cells [88]. Accumulation of intracellular ATP would be anticipated if Panx1 activity was impaired. Indeed, P2RX7-deficient IL-15 polarized CD8 T cells exhibited increased intracellular ATP and a low AMP/ATP ratio which may have contributed to their reduced AMPK activity, and forcing AMPK activation partially restored mitochondrial respiration and memory generation by P2RX7-deficient CD8 T cells [23]. One of the targets of the AMPK pathway is the mTOR signaling complex; the role of P2RX7 in memory CD8 T cell generation appears to rely on this aspect of AMPK function, since negative regulation of the mTOR signaling pathway through low-dose rapamycin can also partially restore memory generation by P2RX7-deficient CD8 T cells [23]. The mTOR complex activation can be negatively modulated by other pathways such as via TSC1/2 activity [89,90]. Whether P2RX7 signaling negatively regulates the mTOR complex through this or other pathways is not yet understood.
As mentioned in the introduction, less is understood about the metabolic requirements for maintenance of already-established memory CD8 T cells. Evidence from previous studies suggest that sustained AMPK activity is important to preserve mitochondrial health of memory CD8 T cells [91]. Likewise, restraint of mTOR activity may ensure sustained fatty acid oxidation (FAO)-mediated mitochondrial metabolism that supports memory CD8 T cell long-term persistence and function [19,20], while high mTOR activity is a hallmark of exhausted, dysfunctional CD8 T cells in response to chronic viral infection [92]. Considering the requirement for P2RX7 for the long-term maintenance of memory CD8 T cells [23] and the aforementioned role of the Panx1-P2RX7 axis in the control of the AMPK-mTOR axis, it is possible that Panx1-mediated ATP release keep the AMP/ATP ratios at a level sufficient to favor AMPK signaling. Consequently, the mTOR signaling pathway is held in check, resulting in homeostatic maintenance of memory CD8 T cells.
A potential decrease in intracellular ATP levels by Panx1 would not be the only cellular metabolic pathway sustained in a counterintuitive way in memory CD8 T cells: circulating memory CD8 T cells rely on de novo synthesized fatty acids for their own fatty acid oxidation (FAO)-mediated increase in mitochondrial respiration [14]. This process is energetically futile since it does not lead to any net gain in intracellular ATP levels. A plausible advantage for that pathway would be guaranteed maintenance of the biomass production machinery together with assurance of mitochondrial health, which would coordinate to provide long-term survival and the ability to quickly mount secondary responses [93]. The proposed role for the Panx1-P2RX7 axis would offer an additional way of guaranteeing overall mitochondrial function and the potential for biogenesis in memory CD8 T cells. Overall, this would evidence the need for controlled energy production in resting memory CD8 T cells, at least in response to acute viral infections.
Based on the knowledge about the metabolic requirements for memory CD8 T cell establishment and long-term maintenance, and about the potential role of eATP sensing for this to happen, we propose a hypothetical model (Figure 1, Table 1) in which, during effector immune responses and the transition to memory, eATP coming primarily from CD8 T cell-extrinsic sources such as dying and/or inflamed cells drive the activation of the P2RX7 pathway. The induced Ca2+ influx leads to activation of the AMPK pathway and subsequent curbing of the mTOR axis function, favoring increased mitochondrial respiration, memory CD8 T cell formation and survival during contraction. Conversely, in the absence of infection or inflammation, CD8 T cell-intrinsic ATP release through Panx1 would predominate, which sustains the eATP ligand for P2RX7 and promotes long-term memory CD8 T cell maintenance. This pathway would support Ca2+ influx through P2RX7 and, at the same time, ATP export through Panx1 may be beneficial for memory CD8 T cells by limiting intracellular ATP. Together, these signals would favor sustained AMPK activation, among other effects. This positive feedback loop (perhaps occurring periodically as circulating cells migrate through the low shear stress environment of lymphoid or non-lymphoid tissues), would favor the long-term survival of memory CD8 T cells in the absence of major cell-extrinsic eATP sources.
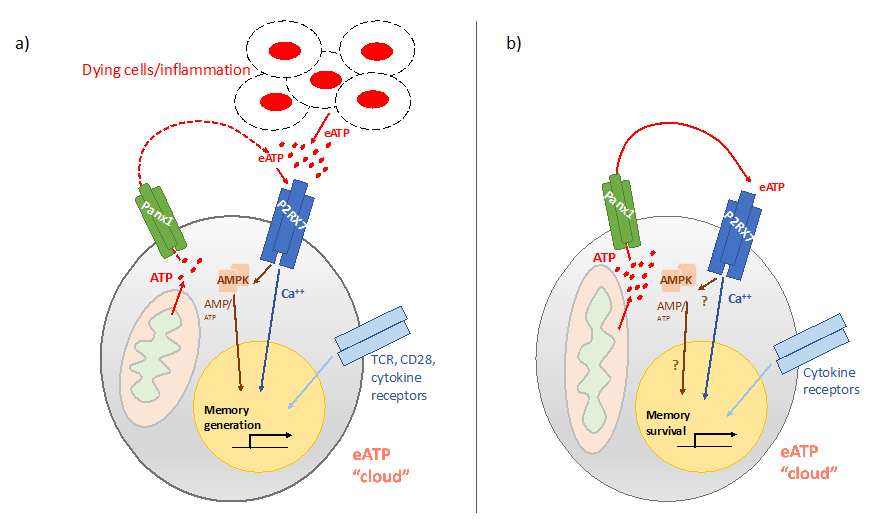 Figure 1. Hypothetic model on the potential role of the Panx1-P2RX7 signaling axis for memory CD8 T cell homeostasis. (a) During effector immune responses or during contraction, CD8 T cell memory precursors express P2RX7, which senses eATP and induces the activation of the AMPK pathway. Together with TCR, costimulatory and cytokine signals, this promotes the generation of memory CD8 T cells. Panx1 is expressed by these cells and likely export ATP, but eATP coming from inflamed/dying cells (due to ongoing infection) is likely dominant. (b) After infection and inflammation is controlled, circulating memory CD8 T cells still rely on P2RX7 for their long-term survival. Cell-extrinsic sources of eATP, however, are expected to be scarce. We hypothesize that Panx1-mediated ATP release is crucial to provide eATP in an autocrine way, thereby guaranteeing the maintenance of a pericellular eATP “cloud”. This maintains the engagement of the P2RX7 signaling pathway. Periodic and/or sustained activation of P2RX7 in memory CD8 T cells could maintain memory CD8 T cells metabolic homeostasis. Simultaneously, ATP export itself maintain the AMP/ATP ratios optimal for AMPK activation, which reinforces the metabolic fitness of memory CD8 T cells through negative regulation of mTOR. Finally, another possible consequence of ATP export is avoiding excessive intracellular ATP levels which would lead to decreased free Mg2+ levels in the cytosol, which is crucial to keep the potential for protein synthesis in memory CD8 T cells. Overall, maintenance of the Panx1-P2RX7 circuit would keep the metabolic homeostasis of memory CD8 T cells and will ensure the ability to rapidly respond to secondary antigen encounters.
Figure 1. Hypothetic model on the potential role of the Panx1-P2RX7 signaling axis for memory CD8 T cell homeostasis. (a) During effector immune responses or during contraction, CD8 T cell memory precursors express P2RX7, which senses eATP and induces the activation of the AMPK pathway. Together with TCR, costimulatory and cytokine signals, this promotes the generation of memory CD8 T cells. Panx1 is expressed by these cells and likely export ATP, but eATP coming from inflamed/dying cells (due to ongoing infection) is likely dominant. (b) After infection and inflammation is controlled, circulating memory CD8 T cells still rely on P2RX7 for their long-term survival. Cell-extrinsic sources of eATP, however, are expected to be scarce. We hypothesize that Panx1-mediated ATP release is crucial to provide eATP in an autocrine way, thereby guaranteeing the maintenance of a pericellular eATP “cloud”. This maintains the engagement of the P2RX7 signaling pathway. Periodic and/or sustained activation of P2RX7 in memory CD8 T cells could maintain memory CD8 T cells metabolic homeostasis. Simultaneously, ATP export itself maintain the AMP/ATP ratios optimal for AMPK activation, which reinforces the metabolic fitness of memory CD8 T cells through negative regulation of mTOR. Finally, another possible consequence of ATP export is avoiding excessive intracellular ATP levels which would lead to decreased free Mg2+ levels in the cytosol, which is crucial to keep the potential for protein synthesis in memory CD8 T cells. Overall, maintenance of the Panx1-P2RX7 circuit would keep the metabolic homeostasis of memory CD8 T cells and will ensure the ability to rapidly respond to secondary antigen encounters.
 Table 1. Key points and hypothesis presented in this Perspective.
Table 1. Key points and hypothesis presented in this Perspective.
Aside from testing the validity of this unified model in the context of acute antigenic exposure, it will be important for future studies to explore how this circuit functions in circumstances where antigen and/or inflammation are not quickly eliminated. For example, during chronic viral infection, CD8 T cell exhaustion is associated with lower levels of mitochondrial respiration and homeostasis [83,94], and presence of an ongoing infection and tissue damage would lead to higher levels of CD8 T cell-extrinsic eATP [38]. We found that P2RX7 is crucial for the establishment and survival of CD8 T cells in this context [23], especially TCF1-expressing “stem-like” cells, which transcriptionally resemble bona fide memory CD8 T cells to some extent [95,96]. It will be important to test the cell-intrinsic role of Panx1 in this context, as well as assess to what extent extrinsic sources of eATP maintain the P2RX7 signaling. Likewise, a recent report showed P2RX7-deficient mice had decreased numbers of tumor-infiltrating CD8 T cells and impaired tumor control [76]. However, more studies with distinct knockout mouse models will be needed to better understand how and when CD8-intrinsic P2RX7 plays a role in CD8 T cell antitumor responses. Memory CD8 T cells residing in non-lymphoid tissues might also be exposed to extrinsic eATP sources. In this context, we found a role for P2RX7 [23], although the magnitude of the role may vary from tissue-to-tissue, which could potentially reflect the degree of exposure to tissue-derived eATP versus autocrine eATP provided by TRM through Panx1. In addition, tissue-resident memory CD8 T cells are metabolically distinct from circulating memory counterparts, particularly those residents in the skin [97], hence the consequences of Panx1-mediated ATP release discussed in this Perspective might not apply. Overall, we believe the potential for memory CD8 T cells of regulate their own metabolism through control of intracellular and extracellular ATP levels may help explain how long-term durability of the memory CD8 T cell pool is consistently achieved in distinct organisms with different levels of acute antigen exposure.
KMW, SCJ and HBdS conceived and wrote the review.
The authors declare that they have no conflicts of interest.
This work was funded by NIH awards AI38903 and AI145147 (SCJ). HBdS is a Paul C. Shiverick/Cancer Research Institute Fellow.
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
41.
42.
43.
44.
45.
46.
47.
48.
49.
50.
51.
52.
53.
54.
55.
56.
57.
58.
59.
60.
61.
62.
63.
64.
65.
66.
67.
68.
69.
70.
71.
72.
73.
74.
75.
76.
77.
78.
79.
80.
81.
82.
83.
84.
85.
86.
87.
88.
89.
90.
91.
92.
93.
94.
95.
96.
97.
Wanhainen K, Jameson SC, Borges da Silva H. Self-Regulation of Memory CD8 T Cell Metabolism through Extracellular ATP Signaling. Immunometabolism. 2019;1:e190009. https://doi.org/10.20900/immunometab20190009

Copyright © 2020 Hapres Co., Ltd. Privacy Policy | Terms and Conditions




