Location: Home >> Detail
Med One. 2019;4:e190015. https://doi.org/10.20900/mo.20190015

Institute of Organismic and Molecular Evolution, Johannes Gutenberg University, 55099 Mainz, Germany
† Present Address: Senckenberg Zentrum für Humangenetik, Weismüllerstraße 50, 60314 Frankfurt am Main, Germany
* Correspondence: David Rosenkranz, Tel.: +49-6131-3924004.
This article belongs to the Virtual Special Issue "Non-coding RNAs in Medicine"
Due to the abundance and conserved role of tRNAs, fragments thereof were considered as mere degradation products for a long time. Lately, however, it was unveiled that these 15–35 nucleotides-long tRNA (-precursor) -derived small RNAs (tsRNAs) can modulate gene expression by different mechanisms and act in a variety of contexts. While some tsRNAs inhibit translation globally by impeding the formation of the translation initiation complex, many studies find tsRNAs to silence target genes in a sequence-specific manner that is potentially mediated by Argonaute proteins. This function plays a role in transposon control, but was also found to be exploited by viruses and trypanosoma to regulate host genes. Beyond their involvement in infectious disease, aberrant tsRNA expression is linked to several other diseases such as cancer or neurological disorders. Furthermore, it was recently shown that tsRNAs residing in sperm of high-fat or low-protein diet mice can act as transgenerational transmitters that induce metabolic disorders and addictive behavior in the offspring. A better understanding of tsRNA-mediated gene regulation pathways, will not only expand our knowledge on how parental lifestyle influences the epigenome of the progeny, but may also enable the development of new drugs and biomarkers.
ncRNA, non-coding RNA; rRNA, ribosomal RNA; tRNA, transfer RNA; nt, nucleotide; miRNA, micro RNA; siRNA, small-interfering RNA; piRNA, PIWI-interacting RNA; RISC, RNA-induced silencing complex; tsRNA, tRNA-derived small RNA; tRH, tRNA half; tiRNA, tRNA-derived, stress-induced small RNA; TOG, terminal oligoguanine; eIF, eukaryotic initiation factor; RG4, RNA G-quadruplex; YBX1, Y-Box Binding Protein 1; mTOG, mini TOG-containing tsRNA; PUS7, Pseudouridine Synthase 7; PABPC1, Polyadenylate Binding Protein 1; MSC, Multisynthetase Complex; RNAi, RNA interference; RSV, respiratory syncytial virus; APOER2, apolipoprotein E receptor 2; PBS, primer binding site; HTLV-1, Human T-Cell Leukemia Virus Type 1; HIV-1, human immunodeficiency virus type 1; LTR, long terminal repeat; ALS, amyotrophic lateral sclerosis; Cyt C, Cytochrome C; JAG2, Jagged Canonical Notch Ligand 2
A major advance in understanding gene expression was the discovery of regulatory non-coding RNAs (ncRNAs). ncRNAs are transcripts that, unlike messenger RNA (mRNAs), do not code for proteins. Apart from mutations within protein-coding genes, perturbations in ncRNA pathways can lead to severe disease.
The most abundant and conserved ncRNAs are ribosomal RNAs (rRNAs) and transfer RNAs (tRNAs). While rRNAs together with ribosomal proteins form the ribosome, the macromolecular machine where protein synthesis takes place, tRNAs translate the codon information of the mRNAs during the protein synthesis as tRNAs with the matching anticodon carry the respective amino acid to the end of the growing protein chain. rRNAs and tRNAs thus constitute central factors in protein translation.
Other ncRNAs such as small ncRNAs (<200 nt) and long ncRNAs (>200 nt) add yet another layer to the control of gene expression, as they can enhance or repress either the transcription (transcriptional level) or the translation (post-transcriptional level) of genes by various mechanism. As their expression is often specific to certain tissues or developmental stages, their regulatory potential critically contributes to cell-specific gene expression. While long ncRNAs can modulate gene expression and protein modifications by acting as a scaffold to bring molecular interaction partners together or to sponge up either antisense mRNAs or other regulators of gene expression, small ncRNAs usually guide effector proteins to their targets. The three most prominent small ncRNA classes, the micro RNAs (miRNAs), the small-interfering RNAs (siRNAs) and the PIWI-interacting RNAs (piRNAs), are loaded onto members of the Argonaute family, which can be subdivided into the AGO and the PIWI clade. While the 26–32 nt long piRNAs interact with proteins of the PIWI clade, the 22–24 nt long miRNAs and 21–22 nt long siRNAs bind to proteins of the AGO clade. Together with the respective Argonaute protein, these small ncRNAs form the heart of the RNA-induced silencing complex (RISC). Depending on the AGO/PIWI paralogue involved, the RISC can act either at the transcriptional level, inducing DNA methylation and histone modifications that lead to changes in chromatin structure, or at the post-transcriptional level by slicing mRNA, inducing its decay via decapping and deadenylation or inhibiting translation.
miRNAs and siRNAs typically post-transcriptionally regulate transcripts that have a target site complementary to the small ncRNA. While siRNAs require almost perfect complementarity to the target mRNA, miRNAs allow more imperfect complementarity for target regulation, requiring near-perfect match only within the 7 nt seed region located in the 5′ portion of the miRNA [1]. Similar to this, the piRNA seed is located at position second to seventh of the 5′ end and effective targeting is established for complementary target sites permitting also GU wobble base pairing [2].
While their ability to regulate gene expression bases on similar principles, the biogenesis of small ncRNAs differs substantially. miRNAs and siRNAs are generated from RNA polymerase II transcribed double-stranded precursors, that get cut by Drosha in the nucleus and are further processed by Dicer in the cytoplasm. The resulting duplex is then taken up by an AGO protein which only keeps the main strand while releasing the so-called passenger strand [3]. piRNAs, however, derive from single-stranded RNA pol II transcripts that mainly originate from conserved genomic loci called piRNA clusters. In a 3′-directed phased manner these precursors are cut by Zucchini that generates the 5′ end of primary piRNAs [4,5]. Pre-mature piRNAs with a bias for a uridine at the 5′ end are loaded on a PIWI protein and get first trimmed and then 2’O-methylated at the 3′ end [6–8]. In case of post-transcriptional silencing, the piRNA guides the PIWI protein to a target site of a transcript, which gets sliced with an offset of 10 nt starting from the 5′ end of the guiding piRNA. This induces the generation of secondary, target-derived piRNAs that are again loaded onto PIWI proteins and in turn trigger the production of piRNAs that resemble primary piRNAs, as they can target and slice complementary piRNA precursors. This results in a self-sustaining so-called ping-pong-loop that, while silencing the target transcripts, generates more piRNAs at the same time [9].
Given their important roles in normal physiology, it is not surprising that the dysregulation of small ncRNA pathways is involved in a variety of diseases. The piRNA pathway was discovered, as its disruption led to a strong infertility phenotype in fruit flies [10]. An active piRNA pathway ensures genomic integrity by silencing transposable elements (so-called selfish, jumping genes). miRNAs, on the other hand, were found to act as oncogenes or tumor suppressors and are potential biomarkers for specific cancer types [11]. Pathologic miRNA expression also plays a role in diabetes and cardiovascular disease [12].
tRNA-Derived Small RNAs as Emerging Small ncRNAsThe recently established high-throughput sequencing techniques of small ncRNA transcriptomes did not only expand the knowledge on the three prominent small ncRNA classes, but also opened up the field to study small ncRNA classes whose functions are far less well understood. One of these emerging new players in the small ncRNA zoo are tRNA-derived small RNAs (tsRNAs). While miRNA, siRNA and piRNA pathways were intensively studied over the last decades, tsRNAs were considered as mere degradation products of the abundant tRNAs until recently. First indications, that tsRNAs have a functional role came from studies in Escherichia coli, where tRNA-halves generated upon bacteriophage T4 infection were found to orchestrate a molecular defense response [13].
In this review, we want to give an overview of how tsRNAs are generated and how they regulate genes, highlighting similarities and differences to other small ncRNA pathways. As tsRNAs were very recently shown to be important players in transgenerational epigenetics of metabolic disorders and addiction behavior, we focus especially on their role in disease.
tRNA precursors are transcribed by RNA polymerase III. Still in the nucleus, the so-called leader and trailer sequences are cut off from the tRNA precursors by the endonucleases RNase P and RNase Z, respectively. The trailer sequence being liberated from the 3′ end of the precursor gives rise to a first class of tsRNAs: the tsRNA1 series (also called 3′ U tsRNAs; Figure 1). tsRNAs of the tsRNA1 series are approximately 15–22 nt in size and have a poly-U-stretch at their 3′ end resembling the Pol-III transcription termination signal [14,15]. If present, intronic sequences are spliced. The mature tRNA sequence is then folded and receives several chemical base modifications. Finally, a CCA signature is attached to its 3′ end before it is exported to the cytoplasm. The structure of tRNAs is highly conserved. With its four arms (D loop, anticodon loop, variable region, TψC loop) and the acceptor stem, the secondary structure of a tRNA resembles a cloverleaf. These secondary structure elements are winded up to an L-shaped tertiary structure. By yet not fully elucidated mechanisms, mature tRNAs can get sliced into tRNA halves (tRHs) and smaller tsRNAs (Figure 1). Please note that within this review the umbrella term tsRNA includes tRNA halves as well as the shorter tRNA-derived small RNAs. Depending from which end of the tRNA the tsRNAs are originating, they are part of the 5′ or the 3′ tsRNA series. Most tRHs are generated by bisecting tRNAs near the anticodon. Smaller tsRNAs can be produced by slicing either mature tRNAs or tRHs. In individual cases of potentially misfolded tRNAs this is done by Dicer [16,17], but generally their production is Dicer-independent [18,19]. tsRNAs seem to have the small RNA typical 5′ terminal phosphate group and 3′ terminal hydroxyl group [14,20]. As 5′ and 3′ tsRNAs derive from mature tRNAs, they likewise carry plenty of base modifications, lack introns and in case of the 3′ tsRNAs (also called 3′ CCA tsRNAs) have the post-transcriptionally added 3′ CCA sequence. Importantly, the pool of tsRNAs within a cell does not at all reflect the abundance of the corresponding paternal tRNAs [15,21]. There is emerging evidence that isodecoder-tRNAs (tRNAs having the same anticodon sequence, but bearing sequence variabilities) are tissue-specifically expressed and that their abundance influences the tsRNA pool generated thereof [22]. Additionally, the cleavage does not seem to occur randomly as certain cleavage sites are enriched for the respective parental tRNAs [15].
Apart from tsRNAs that are expressed under physiological conditions, a different tRH type is produced under stress conditions such as hypoxia. These tRNA-derived, stress-induced small RNAs (tiRNAs; 31–40 nt long) are generated by Angiogenin in mammalian cells (RNY1 in yeast). Angiogenin is a RNase Type A ribonuclease, which under stress conditions preferably slices mature tRNAs within the anticodon loop [23]. This creates 5′ tiRNAs with a 3′ phosphate group and 3′ tiRNAs with an 5′ hydroxyl group [24]. Notably, standard small ncRNA library preparation protocols that rely on the ligation of 5′ and 3′ adapter molecules will necessarily fail to include 5′ tiRNAs, carrying a phosphate group at both ends, and 3′ tiRNAs, carrying a hydroxyl group at both ends. This might have considerably contributed to the fact that these tsRNAs remain under the radar of a special scientific attention.
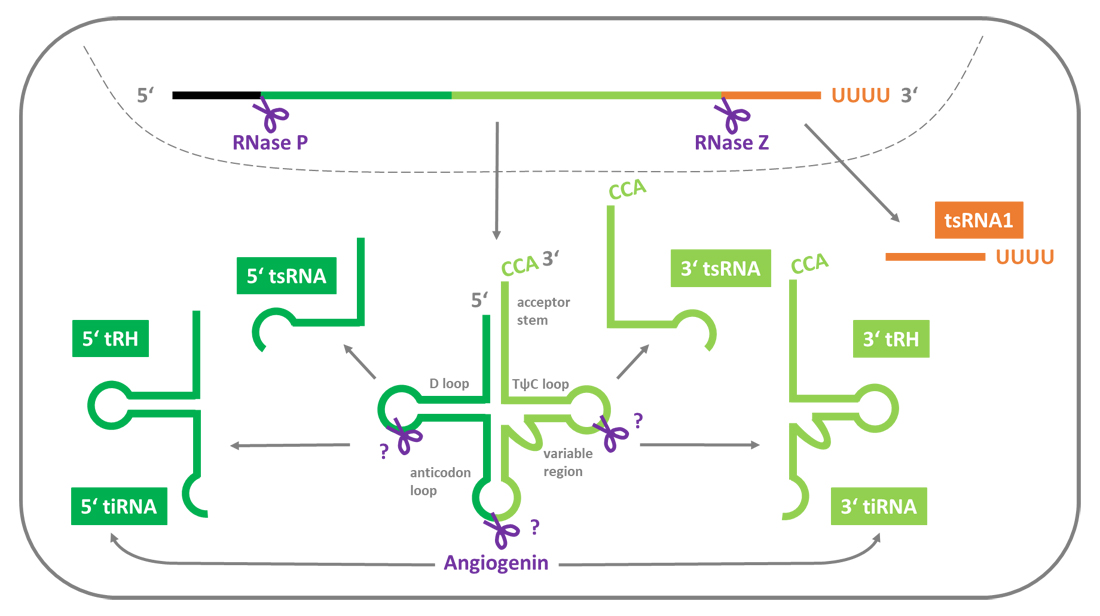 Figure 1. Biogenesis of tRNA-derived small RNAs. The trailer sequences being clipped off from the 3′ end of the tRNA precursors by RNase Z are tsRNAs of the tsRNA1 series (orange). Additionally, RNase P cuts off the leader sequence of the 5′ end of the tRNA precursor. The pre-tRNA is properly folded and extensively post-transcriptionally modified before it gets exported to the cytoplasm. Slicing of mature tRNAs by enzymes such as Angiogenin and Dicer produces tRNA halves and shorter tsRNAs. Depending on their derivation within the parental tRNA, these tsRNAs and tRhs are classified as 5′ tsRNAs/tRHs (dark green) or 3′ tsRNAs/tRHs (light green).
Figure 1. Biogenesis of tRNA-derived small RNAs. The trailer sequences being clipped off from the 3′ end of the tRNA precursors by RNase Z are tsRNAs of the tsRNA1 series (orange). Additionally, RNase P cuts off the leader sequence of the 5′ end of the tRNA precursor. The pre-tRNA is properly folded and extensively post-transcriptionally modified before it gets exported to the cytoplasm. Slicing of mature tRNAs by enzymes such as Angiogenin and Dicer produces tRNA halves and shorter tsRNAs. Depending on their derivation within the parental tRNA, these tsRNAs and tRhs are classified as 5′ tsRNAs/tRHs (dark green) or 3′ tsRNAs/tRHs (light green).
Most tsRNAs are generated from tRNAs that are encoded in the nucleus. However, tsRNAs can also derive from tRNAs that are encoded by organelles such as mitochondria and chloroplasts [25–27]. While in species like Triops cancriformis, nuclear-derived tsRNAs are about 400 times more prevalent than mitochondrial-derived tsRNAs (confirmed by Northern Blot analysis) [27], organellar tsRNAs make up to one quarter of the tsRNA population in Arabidopsis thaliana and are exclusively located in the cytoplasm (confirmed by Northern Blot analysis) [25]. In case of Triops cancriformis, the pool of mitochondrial-derived tsRNAs is more diverse in terms of the region of origin within the parental tRNA, while the nuclear-derived tsRNA pool predominantly consists of 5′ tRHs [27]. Thus, organellar tsRNAs might regulate other biological processes than tsRNAs derived from nuclear encoded tRNAs.
Indications that tsRNAs are generated by defined cutting of tRNAs imply that these small ncRNAs might have a relevant function in the cells. As the major small ncRNA classes are known to regulate gene expression, it seems natural to assume that tsRNAs may also be involved in modulating gene expression. When studying gene regulation mechanisms, an important hint towards the mechanism of action is the subcellular localization of the involved factors. Even though tsRNAs of the tsRNA1 series are generated in the nucleus, they seem to be widely distributed in the cytoplasm (confirmed by Northern Blot analysis) [14,28]. This suggests that they play a role in post-transcriptional gene regulation. 5′ and 3′ tsRNAs are also predominantly localized in the cytoplasm (confirmed by Northern Blot analysis) [14,15]. Contrasting this, 5′ tsRNAs were found to be enriched in the nucleoli deep sequencing data of HeLa cells, suggesting a role in transcriptional regulation, after being reimported to the nucleus [15]. In general, the subcellular distribution of the different tsRNA series is not sufficiently supported by data and requires further investigation.
tsRNAs of all three series could be identified in all domains of life [13,15,29–33]. It is therefore reasonable to assume that they have similar and conserved modes of action in the different organisms. Studies exploring the gene regulatory mechanism of tsRNAs commonly point to two mechanisms: while one cohort of studies describes a sequence-specific post-transcriptional silencing that closely resembles the miRNA pathway, other studies describe a tsRNA-induced global translation repression.
Global Translation RepressionUnder stress conditions, cells globally shut down protein synthesis to quickly adapt their gene expression to the present situation. Part of that is induced by cleaving tRNAs with Angiogenin [23]. This not only decreases the amount of available tRNAs, which slows down translation, but more importantly generates tiRNAs that trigger a stress response program that inhibits the assembly of the translation initiation machinery at mRNAs. Ivanov et al. showed that stress-induced 5′ tiRNAs with a 5′ terminal oligoguanine (TOG; four to five guanine residues) motif such as 5′ tiRNA-Ala displace components of the protein complex eukaryotic initiation factor 4F (eIF4F) from the 5′ cap of mRNAs. This impedes the assembly of translation initiation complexes around mRNAs and in consequence blocks the translation of mRNAs [34]. They could further show that displacing eIF4E and eIF4G is not driven by sequence specificity, but depends on the assembly of four 5′ TOG-containing tiRNAs to a RNA G-quadruplex (RG4) structure (Figure 2A) that might recruit additional factors [35]. This RG4 structure and an intrinsic stem-loop structure of these 5′ tiRNAs are also required for the association with the cold shock domain of Y-Box Binding Protein 1 (YBX1), which induces the formation of stress granules in a phospho-eIF2α-independent manner [36,37]. In these cytoplasmic foci, the mRNAs with attached stalled translation preinitiation complexes are transiently stored and can be released upon stress resolution to quickly reestablish homeostatic translation [38].
Later, Guzzi et al. discovered that 18 nt long TOG-containing tsRNAs, which they refer to as mini TOGs (mTOGs), are enriched in human embryonic stem cells [39]. They could show that only mTOGs that are pseudouridylated at their eighth position (mediated by the Pseudouridine Synthase 7 (PUS7)) repress translation and that their loss impairs early embryogenesis and hematopoietic stem cell differentiation. As they found pseudouridylated mTOGs to strongly interact with Polyadenylate Binding Protein 1 (PABPC1), a central translation initiation factor, while YBX1 predominantly binds to unmodified mTOGs, they hypothesize that pseudouridylation may lead to structural rearrangements that allows mTOGs to orchestrate different cell processes by binding to the respective protein partner.
Similar results were obtained by Krishna et al. [40]. They observed that a specific set of 5′ tRHs (30–35 nt) that is generated independently of Angiogenin is specifically enriched during retinoic acid induced differentiation of mouse embryonic stem cells (confirmed by Northern Blot analysis). As these 5′ tRHs were found to associate with individual subunits of polysomes as well as with completely assembled polysomes this suggests that 5′ tRHs are able to regulate translation at various stages. However, how this is accomplished remains an open question. Further, they could show that depending on the differentiation state these 5′ tRHs are interacting with distinct effector proteins and bind to different mRNA pools suggesting that these 5′ tRHs contribute to stem cell differentiation. As the differentiation-dependent 5′ tRHs mainly interact with the same protein, it is likely that they exert similar functions. Exemplarily for 5′ tRH-Gln-CTG, they provide a mechanistic explanation how these 5′ tRHs drive differentiation: by sequestering the ribosomal binding protein Igf2bp1 the mRNA of cMyc, a potent regulator of pluripotency, is no longer stabilized and gets degraded, which in turn perpetuates differentiation processes.
In case of the model archaeon Haloferax volcanii, protein synthesis is inhibited by binding of a 5′ tsRNA (tsRNA-Val; 26 nt) that is generated under alkaline stress conditions to the small subunit of the ribosome (confirmed by Northern Blot analysis) [29]. The endonuclease that generates this tsRNA is not known. In a follow-up study Gebetsberger et al. introduced synthetic Val-tsRNAs in S. cerevisiae and E. coli, to show that the inhibitory potential of this tsRNA on mRNA translation is conserved in all three domains of life [41]. They further showed that the 5′ tsRNA-Val specifically binds in close proximity to the mRNA channel of the 30S ribosomal subunit. This impedes the binding of this ribosomal subunit to mRNAs and thereby interrupts the formation of translation initiation complexes (Figure 2C).
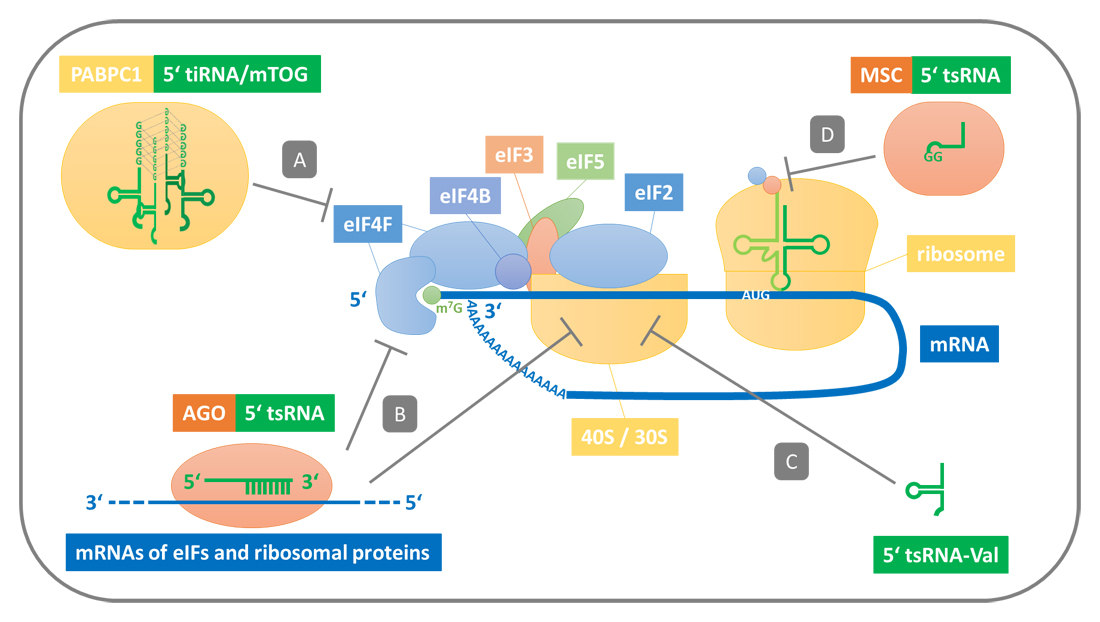 Figure 2. Mechanisms of tsRNA-mediated repression of translation initiation and polypeptide elongation. (A) 5′ tiRNAs with a 5′ TOG motif form a R4G structure that is required to displace the translation initiation complex eIF4F from the 5′ cap of mRNAs. In case of embryonic stem cells this is mediated by the protein PABPC1 that interacts with R4G structures of smaller mTOCs. (B) 5′ tsRNAs associated with Argonaute proteins sequence-specifically silence the mRNAs of translation initiation factors and ribosomal proteins. Target sites are distributed all over the respective mRNAs and efficient silencing is not restricted to a miRNA-like complementarity (see also Figure 3B). (C) 5′ tsRNA-Val binds nearby the mRNA channel of the small ribosomal subunit (eukaryotic 40S and archaeal/bacterial 30S), which impedes binding to mRNA. (D) 5′ tsRNAs with a 3′ terminal GG motif block translation. At least 5′ tsRNA-Gln is primarily associated with the protein complex MSC that potentially inhibits polypeptide elongation.
Figure 2. Mechanisms of tsRNA-mediated repression of translation initiation and polypeptide elongation. (A) 5′ tiRNAs with a 5′ TOG motif form a R4G structure that is required to displace the translation initiation complex eIF4F from the 5′ cap of mRNAs. In case of embryonic stem cells this is mediated by the protein PABPC1 that interacts with R4G structures of smaller mTOCs. (B) 5′ tsRNAs associated with Argonaute proteins sequence-specifically silence the mRNAs of translation initiation factors and ribosomal proteins. Target sites are distributed all over the respective mRNAs and efficient silencing is not restricted to a miRNA-like complementarity (see also Figure 3B). (C) 5′ tsRNA-Val binds nearby the mRNA channel of the small ribosomal subunit (eukaryotic 40S and archaeal/bacterial 30S), which impedes binding to mRNA. (D) 5′ tsRNAs with a 3′ terminal GG motif block translation. At least 5′ tsRNA-Gln is primarily associated with the protein complex MSC that potentially inhibits polypeptide elongation.
With luciferase reporter assays in HeLa cells, Sobala and Hutvagner could show an inhibitory effect on translation for 5′ tsRNAs (e.g., tsRNA-Gln; 19 nt) that are generated independent of Angiogenin (Figure 2D). Here, effective translation inhibition was dependent on a 3′ “GG” dinucleotide sequence in the tsRNA, while a sequence complementarity to the silenced mRNA was not required [42]. As 5′ tsRNAs were found to associate with actively elongating polysomes (confirmed by Northern Blot analysis) and overexpression of 5′ tsRNA-Gln did not reduce the number of reporter mRNAs being present in polysomes, the authors suggest that ribosome assembly is not impaired but the elongation of the peptide chain. By combining isotopic labeling SILAC mass spectrometry analysis with RNA immunoprecipitation, they later identified the Multisynthetase Complex (MSC) as major binding partner of 5′ tsRNA-Gln [43]. As MSC is involved in the processivity of translation, binding of 5′ tsRNA-Gln might impede translation by blocking polypeptide elongation [44–47].
Sequence-Specific Gene RegulationA different mode of translation suppression was revealed by Luo et al. via mRNA sequencing and ribosome profiling of tsRNA mimic transfected S2 cells [48]. Here, 5′ tsRNAs (20–22 nt) do not inhibit translation by displacing or blocking proteins of the translation machinery, but instead silence the mRNAs of key components of the translation machinery (e.g., ribosomal proteins and translation initiation factors) by sequence-specific binding (Figures 2B and 3B). The 7 nt long target sites that were associated with a reduced translation efficiency were perfectly complementary to the respective 5′ tsRNAs and were conserved across different Drosophila genomes. The tsRNA-mediated target silencing was further found to be dependent on AGO2 and the fact that 5′ tsRNAs were co-immunoprecipitated with AGO2 suggests that tsRNAs inhibit specific targets via RNA interference (RNAi). However, unlike miRNAs, these 5′ tsRNAs bind equally effective all over the mRNA and a complementary 5′-portion of the tsRNA is not critical for the silencing effect. A miRNA-untypical gene targeting was also observed by Deng et al. in virus infected A549 cells, where the 3′-portion of a 5′ tRH (tRH-Glu-CTC; 31 nt) suppresses anti-viral target genes by sequence-specific binding to the 3′ UTR of their mRNAs [49]. These findings contradict RNAi-typical Argonaute-induced silencing, as the structural and biophysical environment of the Argonaute MID-PIWI domain favors a tight binding of the 5′-portion of the small RNA, which is then used for target recognition [50].
On the other hand, several studies showed that tsRNAs from different series (18 nt 3′ tsRNA, 5′ and 3′ tsRNAs, 22 nt 3′ tsRNA) not only associate with the different Argonaute paralogs (confirmed by Northern Blot analysis in [18]), but also silence mRNAs via sequence complementary matching of their 5′-portion to target sites within the 3′ UTR (Figure 3A) [15,18,51]. As Kuscu et al. noted, 3′ tsRNA-mediated target silencing mainly depends on a miRNA-like 5′ seed, but sequence complementarity beyond the seed sequence is also required [18]. By performing Renilla luciferase reporter assays in HCT116 cells, Haussecker et al. observed a sequence-dependent target inhibition for 5′ tsRNAs and 3′ tsRNAs, but not tsRNA1s [14]. In line with this, Kumar et al. found 5′ and 3′ tsRNAs, but not tsRNA1s, to associate with the four human AGO paralogs when analyzing PAR-CLIP data from HEK293 cells [15]. While 5′ and 3′ tsRNAs had read counts in the range of miRNAs for AGO1, AGO3 and AGO4, there were almost no read counts for AGO2.
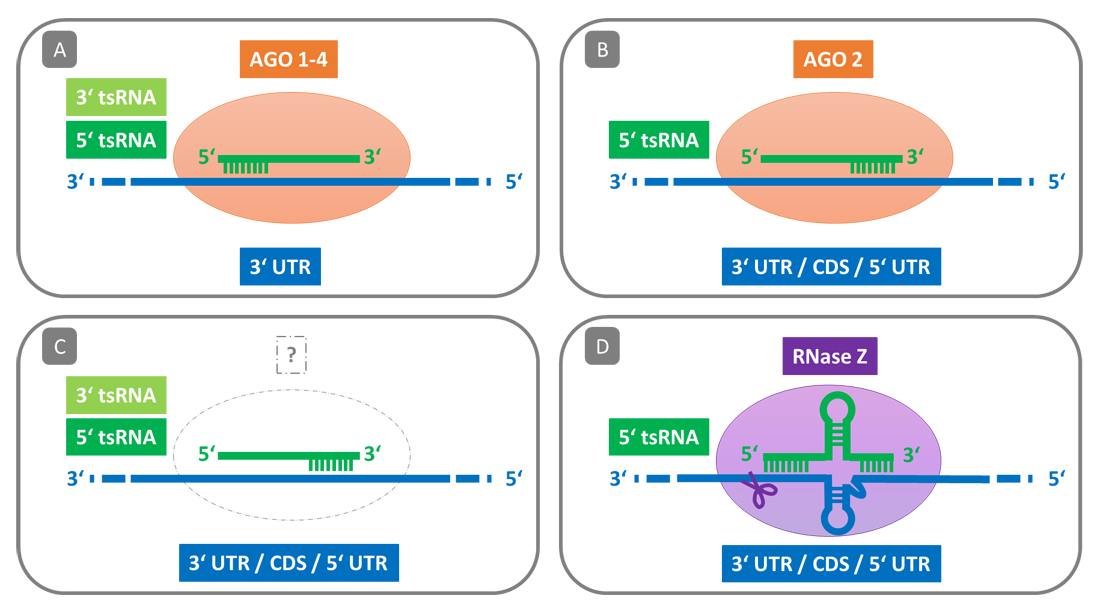 Figure 3. Mechanisms of tsRNA-mediated sequence-specific gene silencing. (A) Typical AGO-mediated silencing: 3′ tsRNAs and 5′tsRNAs, but not tsRNA1s, were found to associate with all four human AGO paralogs. The tsRNA-AGO complexes silence mRNAs that have a target site within the 3′ UTR that is complementary to the 5′ portion of the tsRNA. (B) Untypical AGO2-mediated silencing: 5′ tsRNAs associate with AGO2. Target sites are complementary to various portions of the tsRNA (e.g., the 3′ portion) and distributed all over the mRNA. (C) tsRNAs interact with yet unknown effector proteins or act independent of effectors to silence target mRNAs. Target recognition may be miRNA-untypical. (D) RNase Z mediated silencing: 5′ tRHs can form hybrids with mRNAs that structurally resemble pre-tRNAs. RNase Z recognizes such RNA hybrids and cleaves the mRNA.
Figure 3. Mechanisms of tsRNA-mediated sequence-specific gene silencing. (A) Typical AGO-mediated silencing: 3′ tsRNAs and 5′tsRNAs, but not tsRNA1s, were found to associate with all four human AGO paralogs. The tsRNA-AGO complexes silence mRNAs that have a target site within the 3′ UTR that is complementary to the 5′ portion of the tsRNA. (B) Untypical AGO2-mediated silencing: 5′ tsRNAs associate with AGO2. Target sites are complementary to various portions of the tsRNA (e.g., the 3′ portion) and distributed all over the mRNA. (C) tsRNAs interact with yet unknown effector proteins or act independent of effectors to silence target mRNAs. Target recognition may be miRNA-untypical. (D) RNase Z mediated silencing: 5′ tRHs can form hybrids with mRNAs that structurally resemble pre-tRNAs. RNase Z recognizes such RNA hybrids and cleaves the mRNA.
While Argonaute-mediated RNAi is restricted to eukaryotic cells, tsRNAs are found in all domains of life. It was therefore speculated whether tsRNAs are part of an ancient regulatory pathway that was later complemented by the powerful RNAi pathways in the eukaryotic lineage [52]. If tsRNA-mediated gene regulation is not a eukaryotic-specific trait, this would either require other tsRNA-interacting effector proteins, or regulatory mechanisms that do not rely on tsRNA-interacting protein partners (Figure 3C). One potential tsRNA-directed modulator is the full-length form of the tRNA 3′ processing endoribonuclease tRNase Z, which cuts off the trailer sequence of pre-tRNAs. Due to its highly conserved function, it is expressed in all domains of life. As shown by Elbarbary et al., RNase Z can cut RNA hybrids that have secondary structures like tRNA-precursors. The existence of such RNase Z-interacting small ncRNAs was confirmed by Northern Blot analysis. More importantly, they could show that 5′ tRHs can form such tRNA-precursor-like secondary structures with mRNAs that get recognized by cytoplasmic RNase Z, leading to the endonucleolytic cleavage and thus degradation of these mRNAs (Figure 3D) [53].
Besides these examples of tsRNA-mediated post-transcriptional gene repression, there is also a study by Kim et al. showing that binding of tsRNAs to mRNAs can also have the contrary effect of enhancing translation [54]. They showed that a 3′ tsRNA (tsRNA-Leu-CAG; 22 nt; confirmed by Northern Blot analysis) increases the translation of at least two ribosomal proteins by binding to sequence-complementary target sites that presumably lies within the coding sequence of their mRNA and hypothesize that this binding removes translation-inhibiting secondary structures.
In summary, present studies strongly indicate that tsRNAs can regulate mRNA targets by sequence-specific recognition. However, given the displayed objections, it is still unclear which the effector protein of this tsRNA-mediated gene regulation pathway is and if such an effector is needed in every case. Thus, more research is needed to pin down the exact mechanisms of target regulation.
Increasing evidence is found that environmental stimuli such as an imbalanced diet or stress can influence the gene expression of next generations, which may foster the development of certain disease. This can be mediated by passing on parental small RNAs to the emerging offspring. So far strongest evidence for small RNA mediated transgenerational effects are found in nematodes, plants and ciliates [55,56], but such phenomena could be observed also in mammals [57,58]. First detected for miRNAs, several studies could now show that tsRNAs may also act as transmitters of epigenetic information (Figure 4A). Peng et al. first observed that mature mouse sperm is extremely enriched for 5′ tRHs (29-34 nt) suggesting that these tsRNAs may play a role in early embryo development and transgenerational epigenetic inheritance [59]. Later, Chen et al. could show that male mice subjected to a high-fat diet inherit a susceptibility for metabolic disorders, which is transmitted by 5′ tRHs (30–34 nt) in their sperm [60]. They could trace this back to 5′ tRHs, as microinjection of the 30–40 nt fraction (predominantly 5′ tRHs) from sperm of high-fat diet mice into normal-diet zygotes did phenocopy this effect in the F1 offspring. In a parallel-published study, Sharma et al. aimed to unveil the mechanism of how paternal low-protein diet alters the hepatic cholesterol biosynthesis in the offspring [61]. They could show that sperm of low-protein diet mice contains higher levels of 5′ tRHs (28–34 nt) and indicate that these 5′ tRHs are loaded via vesicles called epididymosomes into the maturing sperm (supported by Northern Blot analysis). Exemplarily for one of the low-protein diet-upregulated tRHs, they show that, in the early embryo, 5′ tRH-Gly-GCC can downregulate the expression of ribosomal proteins as well as transcripts that are driven by the long terminal repeat retrotransposon MERVL. However, it remains unclear whether the repression of MERVL-driven transcripts or the slower growth kinetics due to the impaired ribosomal biogenesis leads to the observed metabolic changes in the offspring of low-protein diet mice.
Chen et al. additionally observed that injecting unmodified tsRNA into zygotes failed to phenocopy the high-fat diet-induced metabolic disturbances, while RNA modifications such as m5C and m2G are enriched in the sperm small RNA population of high-fat diet mice [60]. In a follow-up study, they could show that the tRNA methyltransferase DNMT2 is upregulated in the caput epididymis under high-fat diet conditions [62]. DNMT2 has been shown in previous studies to 5′-methylate the cytosine of certain tRNAs at position 38 [63,64]. By generating Dnmt2-knockout mice, they could demonstrate that DNMT2 is required for intergenerational transmission of high-fat diet-induced metabolic disorders via sperm small RNAs [62]. They showed that DNMT2 elevates the level of m5C modifications in high-fat diet-induced sperm small RNAs and that it alters the composition of the sperm small RNA population (supported by Northern Blot analysis) stressing that RNA modifications are an important feature of small RNA mediated transgenerational effects.
Besides transgenerationally induced metabolic disorders, tsRNAs are also suspected to be a transmitter of inherited behavior. A study from Short et al. suggests that altered levels of miRNAs in mouse sperm, but also altered tsRNA levels (more 5ʹ tRHs, less 3ʹ tsRNAs), are related to inherited affective behaviors, which is reflected by a reduced anxiety and fear memory in the male offspring of physical active mice [65]. A more compelling evidence that sperm tsRNAs also induces behavioral phenotypes across generations was presented by Sarker et al. [66]. Here, they show that feeding pregnant female mice with a high-fat diet led to tsRNA-mediated disturbances of the fat metabolism and addictive-like behaviors in the paternal lineage for at least three generations. Again, microinjection of sperm tsRNAs from F1-males of high-fat diet mice into normal-diet zygotes did phenocopy the observed effects.
Host Regulation by Vesicle-Transferred Parasite tsRNAsAs suggested by Sharma et al., tsRNAs are generated in the epididymis and shuttled via extracellular vesicles into maturing sperm [61]. Another form of vesicle-mediated tsRNA-transfer was observed in the infection process of the parasite Trypanosoma cruzi (T. cruzi). T. cruzi is a parasitic protozoan that has a life cycle with two main developmental stages (replicative or infective) each in an invertebrate and a vertebrate host [67]. Transmission usually occurs when an insect vector (typically members of the Triatominae) feeds on the blood of an infected mammal, thereby taking up the non-replicative trypomastigotes, which are circulating in the bloodstream. Inside insects, the trypomastigotes differentiate into epimastigotes that actively divide before they differentiate into non-replicative metacyclic trypomastigotes. During blood feeding on the next person or animal, the insects release the infective metacyclic trypomastigotes by defecation. Through the bite wound or mucous membranes the parasites can enter the vertebrate host, where they differentiate into replicating amastigotes. After several replication rounds the amastigotes differentiate into bloodstream trypomastigotes that can invade other tissues or can be taken up by new blood feeding insects. Humans infected with this parasite develop the Chagas disease, which is endemic in most parts of Latin America [68].
Although T. cruzi lacks canonical Argonaute-mediated small RNA pathways [69], it can apparently post-transcriptionally regulate gene expression [70]. It was therefore asked whether alternative small RNA pathways have evolved in T. cruzi in order to orchestrate the differentiation processes and adjust gene expression to the changing environment [31]. Thus, several studies analyzed the small RNA transcriptome of T. cruzi and found distinct tsRNA proportions and types in the different developmental forms (confirmed by Northern Blot analysis) [21,31]. While tsRNAs (majorly 5′ tRHs) make up only 26% of the small RNA population in the epimastigote form of T. cruzi, 63% of the small RNAs of the infective metacyclic form are tsRNAs (majorly 3′ tRHs). The developmental stage-dependent tsRNAs not only differ in their abundance and tsRNA class, but also originate from a different set of parental tRNAs and have a distinct subcellular localization pattern (posterior cytoplasmic granules in the epimastigote, even cytoplasmic distribution in the metacyclic form). Later it was revealed that T. cruzi secretes extracellular vesicles that are highly loaded with tsRNAs and TcPIWI-tryp proteins (supported by fluorescence in situ hybridization assays) [71]. TcPIWI-tryp is the only Argonaute protein reported in T. cruzi which was shown to bind tsRNAs [72]. These extracellular vesicles can fuse with other parasites but can also invade mammalian cells. By incorporating extracellular vesicles of T. cruzi into HeLa cells, Garcia-Silva et al. could demonstrate that the tsRNA cargo is able to downregulate HeLa cell genes [73]. As most differentially expressed host genes are related with structural cell components and immune responses pathways, this suggests that vesicle-transmitted tsRNAs play a major role in promoting the susceptibility of the mammalian host cells (Figure 4B). A better understanding of this tsRNA-mediated host-parasite interaction may allow the development of effective anti-parasitic drugs. As trypanosomatids lack orthologues of the mammalian enzymes for tRNA half processing such as Angiogenin [74], targeting Trypanosoma-specific tsRNA biogenesis factors might be a promising approach.
tsRNAs in Virus Infections and RetrotranspositionAnother example of tsRNA-mediated host gene regulation was observed for the respiratory syncytial virus (RSV) [49]. By studying the small RNA population of RSV-infected A549 cells, Wang et al. revealed that virus infection triggers predominantly the production of ~30 nt long 5′ tRHs (confirmed by Northern Blot analysis) [75]. Exemplarily for one of the RSV-induced tsRNAs (5′ tsRNA-Glu-CTC), they could further show that this Angiogenin-dependent tsRNA is able to repress target mRNAs and promote RSV replication (Figure 4B). In a follow-up study, Deng et al. were able to characterize the underlying molecular mechanism [49]. With apolipoprotein E receptor 2 (APOER2) they could not only identify a target gene of 5′ tRH-Glu-CTC, but could also show that this target is an anti-viral protein. Thus, RSV exploits tsRNA-generation of the host to shut down an anti-viral mechanism. APOER2 has a target site for 5′ tRH-Glu-CTC in the 3′ UTR of its mRNA. However, as already indicated previously, they observed a miRNA-untypical target silencing. By performing luciferase assays of target site mutations, they demonstrated that the 3′ end of the 5′ tRH-Glu-CTC is crucial for target repression, while its 5′ end is only contributing to target recognition.
Using 5′ tRHs to modulate host genes might not be a unique mechanism of RSV, as Selitsky et al. observed increased levels of 5′ tRHs in liver samples of human adults that have chronical hepatitis B virus and hepatitis C virus infections [76]. While this study did not investigate whether these 5′ tRHs have a similar role in virus progression, in silico analysis of 5′ tRHs that get produced by Angiogenin upon Rickettsia conorii infection (confirmed by Northern Blot analysis) suggest that endothelial factors of the host are targeted, which would facilitate this bacteria to overcome the hosts endothelial barrier [77].
While the mechanism described for RSV progression bases on a quite specific tsRNA-mediated repression of a host anti-viral protein, tsRNAs may have a more general role in the promotion of retroviruses and retrotransposons, since tRNAs serve as primers for their reverse transcription (Figure 4C) [78]. This may be the case especially for 3′ tsRNAs, as the primer binding site (PBS) is usually complementary to the 3′ end of the priming tRNA. Indeed, Ruggero et al. provided evidence that Human T-Cell Leukemia Virus Type 1 (HTLV-1) exploits host 3′ tsRNAs (18 nt, 3′ tsRNA-Pro) as primer for reverse transcription [79]. When analyzing the small RNA population of CD4+ T cells infected with HTLV-1, they observed that 3′ tsRNA-Pro was the most abundant tsRNA. Previously, its parental tRNA (tRNA-Pro) has been suggested to act as a primer for reverse transcription of HTLV-1 by binding via its 3′ portion to the PBS of HTLV-1 transcripts [80]. In a reverse transcription assay, Ruggero et al. could confirm that 3′ tsRNA-Pro can serve as a primer for HTLV-1 reverse transcriptase [79]. They could further show that both the parental tRNA-Pro as well as the 3′ tsRNA-Pro are enriched in virus particles suggesting an important role for virus progression.
Contrasting conclusions were drawn by Yeung et al., who found high levels of a 18 nt 3′ tsRNA (3′ tsRNA-Lys; confirmed by Northern Blot analysis) in human immunodeficiency virus type 1 (HIV-1) infected MT4 T-cells [81]. 3′ tsRNA-Lys is antisense to the PBS of the HIV-1. As they saw this 3′ tsRNA to be associated with AGO2, they suggested that 3′ tsRNA-Lys directs an RNAi-based defense mechanism of the host against HIV-1. Supporting this hypothesis, they observed a 3′ tsRNA-Lys-dose-dependent depletion of HIV-1 transcripts and further showed that HIV-1 replication is increased in cells where 3′ tsRNA-Lys is antagomir-inhibited.
Similar observations were made for long terminal repeat (LTR)-retrotransposons, for which Schorn et al. could show that 3′ tsRNAs play a role in inhibiting their retrotransposition in mouse stem cells [82]. Retrotransposons are “parasitic genes” within a genome that amplify themselves and jump to other genome loci utilizing a mechanism that involves reverse transcription. Schorn et al. noted that 3′ tsRNAs are especially abundant in mouse stem cells that are depleted for histone H3 lysine 9 tri-methylation and therefore are impaired in transcriptional silencing of retrotransposons. Intriguingly, the 3′ tsRNAs that were found to be enriched are antisense to the 18 nt PBS of the most active mouse transposon families. Using transposition reporter assays, they could not only show that 3′ tsRNAs inhibit retrotransposition, but present two different mechanisms that depend on the size of the 3′ tsRNA. While 18 nt 3′ tsRNAs are only able to block the reverse transcription by binding to the PBS of LTR-retrotransposon transcripts, 22 nt 3′ tsRNAs post-transcriptionally silence the LTR-retrotransposons by reducing the retrotransposon transcript levels. The authors suggest that this effect is mediated by an RNAi-mechanism, as in a previous study 22 nt 3′ tsRNAs were shown to cleave target RNA in association with AGO2 [19].
Instead of complementarity to the 3′ portion of tRNAs, few retrotransposons have a PBS that is complementary to an internal region of the primer tRNA. In case of the fruit fly retrotransposon copia, which is such an exceptions, it was suggested that the 39 nt 5′ tRH-Met, but not its parental tRNA, serves as a primer for reverse transcription [78]. Similar to the situation in retroviruses, 5′ tRHs may not only promote retrotransposon activity, but may also inhibit it. As described above, 5′ tRHs transmitted via sperm (confirmed by Northern Blot analysis) are able to downregulate transcripts that are driven by the LTR-retrotransposon MERVL in the developing mouse embryo [61]. Other indications come from plants, where 19 nt 5′ tsRNAs that are enriched in the pollen of Arabidopsis (confirmed by Northern Blot analysis) associate with AGO1 and cleave LTR-retrotransposons of the gypsy family [83].
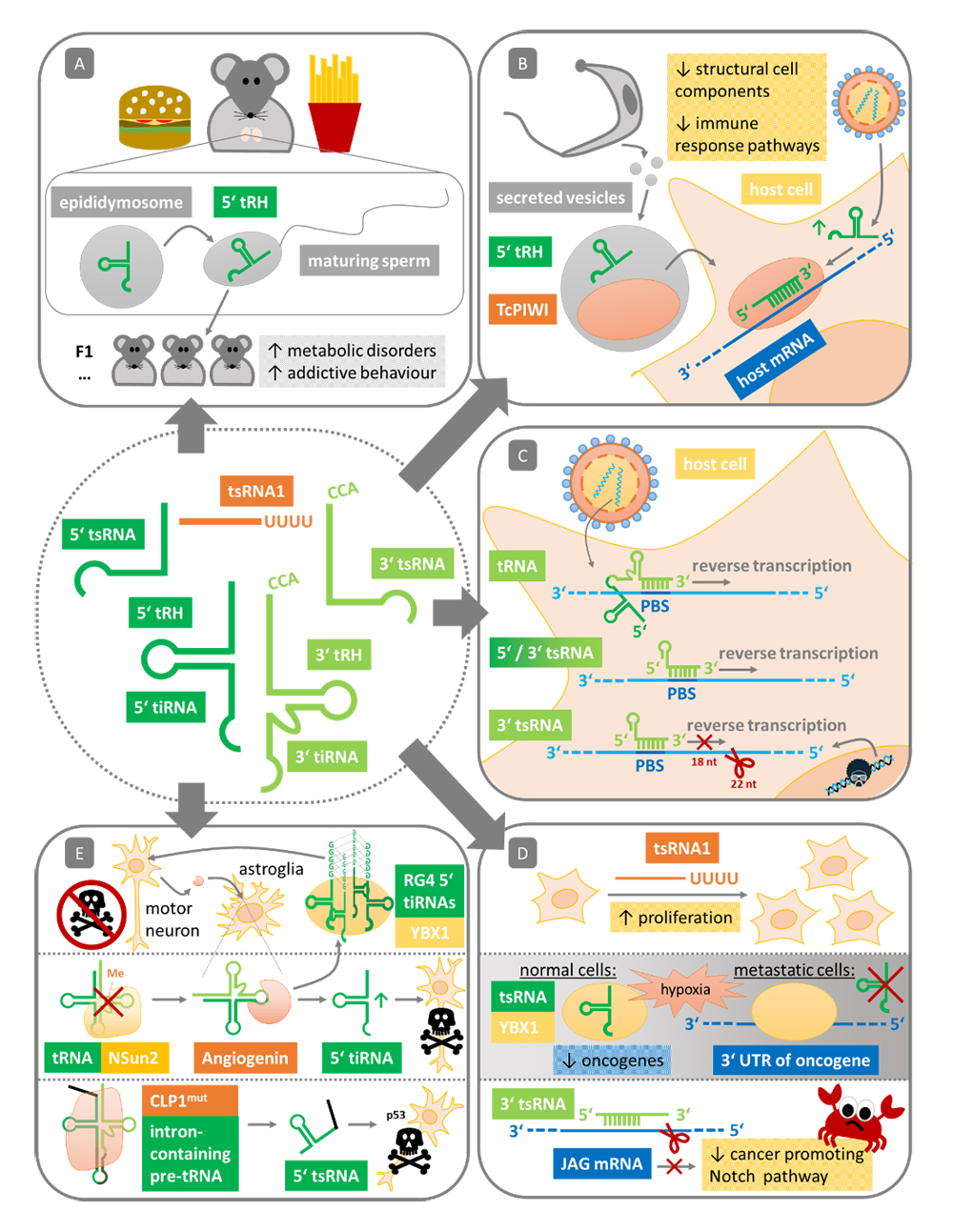 Figure 4. Association of tsRNAs with diseases. (A) 5′ tRHs as transgenerational transmitters inducing metabolic disorders and addictive behavior. 5′ tRHs are loaded into maturing sperm via epididymosomes. (B) tsRNAs repress host genes to facilitate infection. T. cruzi secretes vesicles containing tRHs and TcPIWI-tryp proteins that can fuse with host cells. RSV infection triggers the accumulation of host-generated 5′ tRHs. (C) Generally, the 3′ portion of tRNAs primes reverse transcription of retroviruses. Similarly, tsRNAs can act as primers, but can also block primer binding sites (PBS) of RNA from retroviruses and retrotransposons. (D) Proliferation-enhancing tsRNA1s can foster cancer progression (upper part), while, e.g., specific 3′ tsRNAs act anti-tumorigenic. tsRNAs generated under hypoxia sequester YBX1, which prevents stabilization of pro-oncogenic transcripts. As metastatic cells inhibit generation of these tsRNAs, YBX1 is able to stabilize tumor-promoting mRNAs (middle part). By degrading the mRNA of JAG, a 3′ tsRNA inactivates the cancer promoting Notch pathway (lower part). (E) Angiogenin secreted by stressed motor neurons is endocytosed by adjacent astroglia where tRNAs are cleaved to generate tiRNAs. 5′ tiRNAs forming R4G structures can associate with YBX1, which triggers stress granule assembly that contributes protecting motor neurons from undergoing apoptosis (upper part). Angiogenin-mediated tiRNA-generation can also promote apoptosis when the tRNA-methylase NSun2 is defective (middle part). Mutations in the Kinase CLP1 induce the accumulation of leader-sequence-containing 5′ tsRNAs, which trigger p53-dependent apoptosis of motor neurons (lower part).
Figure 4. Association of tsRNAs with diseases. (A) 5′ tRHs as transgenerational transmitters inducing metabolic disorders and addictive behavior. 5′ tRHs are loaded into maturing sperm via epididymosomes. (B) tsRNAs repress host genes to facilitate infection. T. cruzi secretes vesicles containing tRHs and TcPIWI-tryp proteins that can fuse with host cells. RSV infection triggers the accumulation of host-generated 5′ tRHs. (C) Generally, the 3′ portion of tRNAs primes reverse transcription of retroviruses. Similarly, tsRNAs can act as primers, but can also block primer binding sites (PBS) of RNA from retroviruses and retrotransposons. (D) Proliferation-enhancing tsRNA1s can foster cancer progression (upper part), while, e.g., specific 3′ tsRNAs act anti-tumorigenic. tsRNAs generated under hypoxia sequester YBX1, which prevents stabilization of pro-oncogenic transcripts. As metastatic cells inhibit generation of these tsRNAs, YBX1 is able to stabilize tumor-promoting mRNAs (middle part). By degrading the mRNA of JAG, a 3′ tsRNA inactivates the cancer promoting Notch pathway (lower part). (E) Angiogenin secreted by stressed motor neurons is endocytosed by adjacent astroglia where tRNAs are cleaved to generate tiRNAs. 5′ tiRNAs forming R4G structures can associate with YBX1, which triggers stress granule assembly that contributes protecting motor neurons from undergoing apoptosis (upper part). Angiogenin-mediated tiRNA-generation can also promote apoptosis when the tRNA-methylase NSun2 is defective (middle part). Mutations in the Kinase CLP1 induce the accumulation of leader-sequence-containing 5′ tsRNAs, which trigger p53-dependent apoptosis of motor neurons (lower part).
In conclusion, tsRNAs can be accounted to the pool of small ncRNAs that repress reactivated retrotransposons, thereby safeguarding the genome of germline cells and stem cells. While a tsRNA was also shown to inhibit virus progression by degrading viral transcripts, several studies found retroviruses to exploit tsRNAs to foster their own replication, or to downregulate the host’s defense mechanisms. Targeting infection-promoting tsRNAs may therefore be a promising attempt to develop anti-viral treatment.
tsRNAs in Cancer ProgressionInitially, tsRNAs were deeper characterized and recognized as functional small ncRNAs in the context of cancer. In 2009 Lee et al. analyzed small RNA sequencing data of the human prostate cancer cell line HCT116 [84]. Here, they found tsRNAs to be the second most abundant small ncRNA class following miRNAs. They deeper investigated a tsRNA1 (tsRNA1-Ser-TGA, 19 nt), which was more abundant in small RNA libraries of a variety of cancer cell lines compared to normal tissue libraries (confirmed by Northern Blot analysis). By siRNA-mediated knockdown and rescue co-transfection, they could show that tsRNA1-Ser-TGA is required for cell cycle propagation and proliferation of HCT116 cells (Figure 4D, upper part). As luciferase reporter assays did not reveal any modulation effect, the authors suggest an RNAi-independent mechanism, which is in line with the above-mentioned PAR-Clip data analysis of Kumar et al. [15].
So far, the best mechanistic explanation for a role of tsRNAs in cancer progression was found for a specific set of hypoxia-induced tsRNAs (Figure 4D, middle part) [85]. As hypoxia is a major stress encountered by progressing cancerous cells, Goodarzi et al. analyzed the tsRNA levels in breast cancer cells under low oxygen conditions. While hypoxia induced the expression of a group of tsRNAs in breast cancer cells (MDA-231 cells) as well as in normal mammary epithelial cells, this specific set of tsRNAs was not upregulated in highly metastatic breast cancer cells (MDA-LM2 cells), indicating that these tsRNAs might have a tumor suppressive role. Interestingly, these particular tsRNAs (tsRNA-Glu, -Asp, -Gly, and -Tyr) share a sequence motif (“SCUBYC”) suggesting that they interact with a common trans factor. Indeed, by using a streptavidin-coupled tsRNA-Gly mimetic, they could identify the RNA-binding protein YBX1 as a specific interactor. As mentioned previously, YBX1 was found to induce stress granule formation when interacting with 5′ tiRNAs [34]. Unlike the 5′ tiRNAs of the study conducted by Ivanov et al., the tsRNAs here are not generated by a cleavage in the anticodon loop of the parental tRNA [34], but originate from a cleavage site within the D loop and cover at least the complete anticodon loop [85]. High-throughput sequencing of the YBX1-crosslinked small ncRNAs confirmed a specific interaction with this subset of tsRNAs. The fact that relatively low expressed tsRNAs (tsRNA-Glu and -Asp) were found to be enriched in this YBX1-crosslink sequencing data, but not highly expressed tsRNAs that are not part of this potentially tumor suppressive tsRNA set, suggests that the relative abundance of tsRNAs not necessarily represents an indicator for functional activity. Goodarzi et al. could further show that this specific set of hypoxia-induced tsRNAs competes with pro-oncogenic transcripts for YBX1 binding. As YBX1 stabilizes pro-oncogenic transcripts by binding to their 3′ UTR, displacement of YBX1 from these mRNAs leads to their degradation, which in turn inhibits tumor progression. In contrast, highly metastatic breast cancer cells evade this mechanism, as they are able to block the induction of these tumor suppressive tsRNAs. Here, YBX1 continues to stabilize pro-oncogenic transcripts, which is fostering metastasis of these breast cancer cells even more.
Another tumor suppressive tsRNA was found to be involved in colorectal cancer [86]. Since this specific 17 nt 3′ tsRNA can be derived from both, the 3′ end of tRNA-Leu, as well as from the pre-miRNA-1280 (supported by Northern Blot analysis), 3′ tsRNA-Leu is also listed as miR-1280 in popular databases. Several studies indicated a tumor suppressive activity of 3′ tsRNA-Leu/miR-1280 in various cancer types such as melanoma, medulloblastoma, bladder cancer and thyroid carcinoma [87–92]. In line with this, Huang et al. demonstrated that 3′ tsRNA-Leu/miR-1280 inhibits the proliferation and colony formation of colorectal cancer cells (Figure 4D, lower part) [86]. They could mechanistically explain this as they identified Jagged Canonical Notch Ligand 2 (JAG2) as a direct target of 3′ tsRNA-Leu/miR-1280. Upon binding of 3′ tsRNA-Leu/miR-1280 to the 3′ UTR of JAG2, the JAG2 transcript gets degraded, which in turn inactivates the Notch signaling pathway. Notch signaling is important for key functions of cancer progression such as proliferation, metastasis and the maintenance of cancer stem cell phenotypes. 3′ tsRNA-Leu/miR-1280 can therefore be considered as a powerful tumor suppressive factor.
In contrast to that, a slightly bigger tsRNA originating from the 3′ end of tRNA-Leu (tsRNA-Leu-CAG; 22 nt; confirmed by Northern Blot analysis) was shown to be essential for viability of the cancer cell lines HCT116 and HeLa [54]. As inhibition of this 3′ tsRNA in a hepatocellular carcinoma mouse model induced apoptosis of the tumorigenic cells, 3′ tsRNA-Leu-CAG might be used as a potential drug target. The fact that two similar tsRNAs may induce completely converse functions is astonishing, but may be a prevalent feature of tsRNA-mediated processes in tumor progression. In line with this are the results of a study that compared the small RNA profiles of different prostate cancer states. While samples from organ-confined prostate cancer predominantly had 18 nt long tsRNAs, samples from metastatic lymph node prostate cancer expressed mainly 27 nt long tsRNAs [93]. It is therefore likely that specific tsRNA populations play different roles in tumor progression.
Beside these reports on the implication of individual tsRNAs in cancer progression, tiRNAs generated by Angiogenin may play a more general role, as this ribonuclease is upregulated in almost all types of cancer [94]. This assumption is supported by the finding, that the ribonucleolylitc activity of Angiogenin is required for angiogenesis and thus tumor formation [95,96]. How exactly tiRNAs are involved in cancer progression is not clear yet.
tsRNAs in NeurodegenerationWhile its relevance for cancer progression is yet unclear, several studies could demonstrate that Angiogenin treatment is able to protect stressed motor neurons from degeneration [97–100]. Mutations of Angiogenin are strongly associated with amyotrophic lateral sclerosis (ALS) and Parkinson disease, where the degeneration of motor neurons is a major symptom [101,102]. Notably, most of the mutations that are associated with ALS impair the RNase function of Angiogenin [102,103]. Thus, it was suggested that Angiogenin-generated tiRNAs play an important role in the survival of motor neurons. Skorupa et al. revealed that stressed motor neurons secrete Angiogenin, which is endocytosed by astroglia in close proximity [104]. Astroglia that are deficient in Angiogenin under normal conditions then generate tiRNAs, which seem to mediate neuroprotection of motor neurons in paracrine. Ivanov et al. could later demonstrate that 5′ tiRNA-Ala and 5′ tiRNA-Cys forming RG4 structures via their 5′ TOG motif can enter motor neurons, where they interact with YBX1 to induce stress granule formation, which might be a central part of the neuroprotective response (Figure 4E, upper part) [37]. As the pathological GGGGCC repeats in the C9ORF72 gene, which is the most common genetic cause of ALS [105], are able to form similar RG4 structures, Ivanov et al. suggest that these repeats interfere with the tiRNA-YBX1-mediated processes such as stress granule formation that in turn induces apoptosis [37].
Angiogenin-treatment was also shown to protect cortical neurons that are exposed to hyperosmotic stress [106]. Even though not validated for the neurons, the study by Saikia et al. provided a mechanism for cell survival of similarly stressed mouse embryonic fibroblasts. They showed that a set of 20 Angiogenin-dependent tiRNAs can form complexes with Cytochrome C (Cyt C) that gets released by the mitochondria in order to trigger the apoptosis cascade. The tiRNA-Cyt C interaction, however, blocks the formation of apoptosomes, which prevents that cells undergo apoptosis.
Contrasting effects were observed by Blanco et al., where accumulation of Angiogenin-generated 5′ tiRNAs in NSun2-mutants leads to reduced cell size and increased apoptosis of cortical, hippocampal and striatal neurons (Figure 4E, middle part) [107]. NSun2 is a cytosine-5 RNA methyltransferase, whose mutation can cause hereditary intellectual disability and the microcephaly causing Dubowitz-like syndrome [108–110]. Blanco et al. could demonstrate that functional NSun2 methylates all tRNAs carrying a cytosine in their variable loop at position 48 or 49 and that this m5C modification protects the tRNA to be cleaved by Angiogenin [107]. In NSun2-deficient cells, target tRNAs remain hypo-methylated and are cleaved by Angiogenin, which leads to an accumulation of 5′ tiRNAs. As 5′ tiRNAs are able to block translation globally [34], the lower translation rates may explain the reduction of neuronal cell size and synapse formation in NSun2 knockout brains [107]. Importantly, these neurodegenerative phenotypes can be reverted by inhibiting Angiogenin.
Another report where tsRNAs are involved in degeneration of motor neurons links distortions in tRNA biogenesis to neurological defects in the peripheral and the central nervous system [111–113]. Hanada et al. showed that mutations in the RNA kinase CLP1 leads to defects in the processing of intron-containing pre-tRNAs, which gives rise to untypical 5′ tsRNAs that contain the 5′ leader sequence (confirmed by Northern Blot analysis) [111]. Accumulation of these 5′ leader exon tsRNAs in motor neurons can trigger the activation of p53-dependent apoptosis (Figure 4E, lower part). As shown by Schaffer et al. these neurodegeneration effects of mutant CLP1 is a conserved feature in vertebrate neurogenesis [113].
Given the broad range of known tsRNA-associated diseases, it is likely that more diseases will be linked to aberrant tsRNA expression in the near future. To be able to develop tsRNA-based biomarkers or therapies, a deeper understanding of tsRNA pathways is required. Major remaining questions are:
•
•
•
•
•
While answering these questions, future studies should consider the following aspects. First of all, in most of the reviewed studies conclusions are drawn from experiments that focus on individual tsRNAs. However, as we learn from the overall picture these studies give, different subsets of tsRNAs even from the same biogenesis-pathway may exert different functions in distinct contexts. It is therefore essential to investigate the mechanisms of different tsRNA subpopulations conjunctively within one experimental setting. Furthermore, most of the studies test the regulatory potential of tsRNAs using synthetic mimetics. As tsRNAs are heavily modified and it was shown that RNA modifications can be important features of small RNA function, the use of mimetics without the endogenous base modifications might lead to artifact results and wrong conclusions. It will be interesting to see whether mimetics including the natural base modifications will have, e.g., different affinities to certain protein interactors. In line with this, future studies should investigate differences in post-transcriptional modifications of tsRNAs and their parental tRNAs. This may not only explain why specific tsRNAs are enriched compared to their source tRNA, but also reveal why certain tsRNAs have different effects under distinct conditions. It is likely that aberrant tsRNA modifications are a major feature in the development and progression of disease. Moreover, future studies should also investigate the role of organelle-derived tsRNAs. This might be especially relevant as mitochondrial-derived tsRNAs can be differentially expressed during development and as RNA modifications within mitochondrial tRNAs were shown to be associated with mitochondrial disease [27,114]. Additionally, library preparation protocols should be improved in order to obtain unbiased sequencing data. Currently, post-transcriptional base modifications as well as untypical 5′ end 3′ end modifications (e.g., tiRNAs) are factors that prevent tsRNAs from being faithfully represented in RNA sequence libraries and can cause sequencing arrests of longer transcripts that would then be erroneously interpreted as tsRNAs. Thus, the development of more sophisticated adapter ligation and reverse transcription protocols is required to obtain an unbiased picture of the real tsRNA population within biological samples. In addition, it advisable to verify small RNA-sequencing based expression data with Northern Blot analysis. Taking these considerations into account, research in this emerging field will open possibilities to develop therapies against certain viruses or parasites or to improve the diagnosis of severe disease such as cancers or neurodegenerative disorders.
The manuscript was written by JJ and DR. Figures were designed by JJ.
The authors declare that there is no conflict of interest.
This work was supported by the International PhD Programme (IPP) coordinated by the Institute of Molecular Biology IMB, Mainz, Germany, funded by the Boehringer Ingelheim Foundation.
We thank Daniel Gebert, Katharina Mauer, Sacha Heerschop, and Hans Zischler for helpful comments and discussion.
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
41.
42.
43.
44.
45.
46.
47.
48.
49.
50.
51.
52.
53.
54.
55.
56.
57.
58.
59.
60.
61.
62.
63.
64.
65.
66.
67.
68.
69.
70.
71.
72.
73.
74.
75.
76.
77.
78.
79.
80.
81.
82.
83.
84.
85.
86.
87.
88.
89.
90.
91.
92.
93.
94.
95.
96.
97.
98.
99.
100.
101.
102.
103.
104.
105.
106.
107.
108.
109.
110.
111.
112.
113.
114.
Jehn J, Rosenkranz D. tRNA-Derived Small RNAs: The Good, the Bad and the Ugly. Med One. 2019;4:e190015. https://doi.org/10.20900/mo.20190015

Copyright © 2020 Hapres Co., Ltd. Privacy Policy | Terms and Conditions




