Location: Home >> Detail
Med One. 2019;4:e190019. https://doi.org/10.20900/mo.20190019
 ,
Youchao Jia 1,*
,
Youchao Jia 1,*
1 Hebei Key Laboratory of Cancer Radiotherapy and Chemotherapy, Department of Medical Oncology, Affiliated Hospital of Hebei University, Baoding 071000, China
2 Department of Chemistry & Biochemistry, Texas State University, San Macros, TX 78666, USA
* Correspondence: Zhenze Zhao; Youchao Jia.
Small cell lung cancer (SCLC) is an invasive and high-grade neuroendocrine malignant tumor. It is characterized by short doubling time, high proliferation rate, and early extensive metastasis. SCLC is sensitive to radiotherapy and chemotherapy in the initial stage; however, it can easily relapse and develop drug resistance. In the past five years, there has been a resurgence of research on SCLC worldwide, including the establishment of SCLC cells, the development of related genetically engineered mouse models (GEMMs) and the establishment of patient-derived xenograft models (PDXs). These studies have identified new potential therapeutic vulnerabilities for SCLC, leading to new clinical trials. In this perspective, the establishment, application, and advantages and disadvantages of three preclinical research models of SCLC are systematically summarized.
Small cell lung cancer (SCLC) is an invasive and highly malignant neuroendocrine tumor with predilection for rapid growth, high early metastatic potential, acquired therapeutic resistance and poor clinical prognosis. In 1926, SCLC was described as a lung tumor called “oat cell sarcoma”. There have been more than 30 years of clinical trials aiming at improving the treatment of SCLC. However, only a small percentage of patients with localized early-stage disease could be cured with surgery or concomitant chemotherapy and radiation. The standard chemotherapy consists of platinum agents (cisplatin or carboplatin) combined with etoposide. In a recent large randomized clinical trial, the median overall survival time of patients with metastatic SCLC receiving standard chemotherapy was 9–11 months [1–4]. Anti-programmed death-ligand 1 antibody atezolizumab was recently added to the first-line therapy for advanced SCLC. However, studies showed less than 15% of patients remained progression-free at one year. The improved median survival for advanced SCLC ranged from 10.3 to 12.3 months [5]. Clearly, more effective regimens are needed for SCLC therapy.
In 1970, Shimosato Y. et al. of Tokyo National Cancer Center extracted SCLC cells from metastatic lymph nodes of oat cell carcinoma and established a human lung cancer cell line. The cell line is named “OAT cell line” due to oat cell carcinoma [6]. The established SCLC cell lines were composed of loose or tightly aggregated floating cells, which grew freely in the culture medium. From 1976 to 1991, the Dartmouth group successfully cultured 122 SCLC cell lines, known as “Dartmouth cell lines” [7]. In the initial stage, the selection of culture medium was a difficult point, as the conventional RPMI-1640 medium containing fetal bovine serum could not meet the requirements of the sustainable selective growth of SCLC cells. In subsequent studies, a specific serum-free medium was developed by Elizabeth Simms. Then, five supplements (hydrocortisone, insulin, transferrin, estradiol, and selenium, HITES) were identified and added to support the continuous replication of established SCLC cells, which was hence known as “HITES medium” [8]. After the SCLC cells are established, researchers usually use RPMI-1640 medium containing fetal bovine serum or completely serum-free limited medium to maintain the established cell lines [9].
From the study conducted by the National Cancer Institute of the United States [10], considerable heterogeneity was found in the morphology of 50 SCLC cell lines. Most cell lines (46 of 50, 92%) grew into dense to loosely arranged suspended aggregates (Types 1–3), while three cell lines (6%) grew into adherent monolayer cultures (Type 4). One of the 50 SCLC cell lines showed a mixed form of Types 2 and 4. Based on the expression of four biomarkers, SCLC cell lines can be divided into two categories. One category is the classical SCLC cell line with increased expression of l-dopa decarboxylase (DDC), bombesin-like immunoreactivity (BLI), neuron-specific enolase (NSE) and creatine kinase brain isozyme (CK-BB). The other is a mutant SCLC cell line lacking key amine precursor uptake and decarboxylation enzymes DDC and CK-BB. There are variants of SCLC cells that do not express BLI, but still have an increased expression of NSE and CK-BB.
Using immortalized human bronchial epithelial cells as a control, researcher can determine the genetic and epigenetic differences between tumor cells and normal cells to study the pathogenesis of SCLC and to screen and test of novel chemotherapeutic drugs [11,12]. Based on gene expression profiling, Rudin et al. recently proposed a new model of SCLC subtypes defined by differential expression of four key transcription regulators: achaete-scute homolog 1 (ASCL1), neurogenic differentiation factor 1 (NeuroD1), yes-associated protein 1 (YAP1) and POU class 2 homeobox 3 (POU2F3). Most SCLCs are either ASCL1-high or NeuroD1-high neuroendocrine subtypes. A small fraction of SCLC is ASCL1-low and NeuroD1-low. These tumors lack neuroendocrine markers and appear to fall into YAP1-high or POU2F3-high subtypes [13].
There are still many limitations in the current establishment of SCLC cell lines. Firstly, although these cells retain driving proto-oncogenes, the lack of interaction with tumor microenvironment may limit the study of immunotherapy or vascular targeted therapy. Secondly, although they are very helpful for in vitro experiments, e.g., drug screening and targeted gene therapy testing, the application of those screening or testing results for in vivo study needs further evaluation. Thirdly, most of the cell lines were cultured in two dimensions, and the cultured SCLC cells maintained the appearance and differentiation characteristics of SCLC cytology, but the ability of differentiation after inoculation into the animal model remained to be observed. Therefore, although the establishment of SCLC cell lines has accelerated the research process of SCLC, there are still problems in many aspects.
Although a cell line is not the ideal model system, it complements the disadvantage of tumor tissue and animal models in studying SCLC. The establishment of SCLC cell line has accelerated the research process of SCLC. Many variant lines confirmed the amplification of MYC gene family in SCLC and the important role of hedgehog pathway and Notch pathway in SCLC [14–17]. At the same time, the SCLC cell lines provide a suitable model for genome (high throughput DNA sequencing) [18], transcriptome (microarray analysis) [19,20], methylation (detection of whole genome methylation sequence) [21] and microRNA analyses [22]. Since the established SCLC cell lines retain the original neuroendocrine phenotype of SCLC cells, they provide a good tool for studying SCLC heterogeneity, transformation, and epigenetics.
Mouse models are widely used in cancer therapeutics to evaluate the efficacy of novel drug candidates in vivo. Historically, these models mainly relied on xenograft systems that use human tumor cell lines and immunodeficient mice to obtain xenograft tumors [23]. However, the results obtained by these models cannot fully predict the clinical outcome and prognosis of SCLC [24]. Therefore, it is very important to find more preclinical models of SCLC to develop new and effective treatment schemes. Several genetic engineering mouse models were developed in recent years.
It was found that the inactivation rate of Rb1 and Tp53 in SCLC patients was as high as 90% [25,26]. Based on this finding, Berns et al. first developed a GEMM for SCLC in 2003 [27]. They introduced loxP sites into key exons of Tp53 and Rb1 tumor suppressors genes and used intratracheal delivery of CRE recombinase by adenovirus infection to inactivate both Tp53 and Rb1 in respiratory epithelium cells in mice. This double-gene-knockout model well simulates the histologic types and metastasis patterns of SCLC. The use of adenoviruses with cell-type specific calcitonin or surfactant protein C promoters in this model indicated that SCLC may arise from at least two different cells of origin: efficient tumorigenesis from lung epithelial cells and less efficient tumorigenesis from alveolar type 2 cells. Then, the researchers developed a series of triple-knockout-variant models to shorten the long incubation period of the double-gene-knockout model. However, the multiple variations of the triple-knockout-models often lead to greater morphological heterogeneity of the resulting tumor. Table 1 summarizes several representative GEMMs.
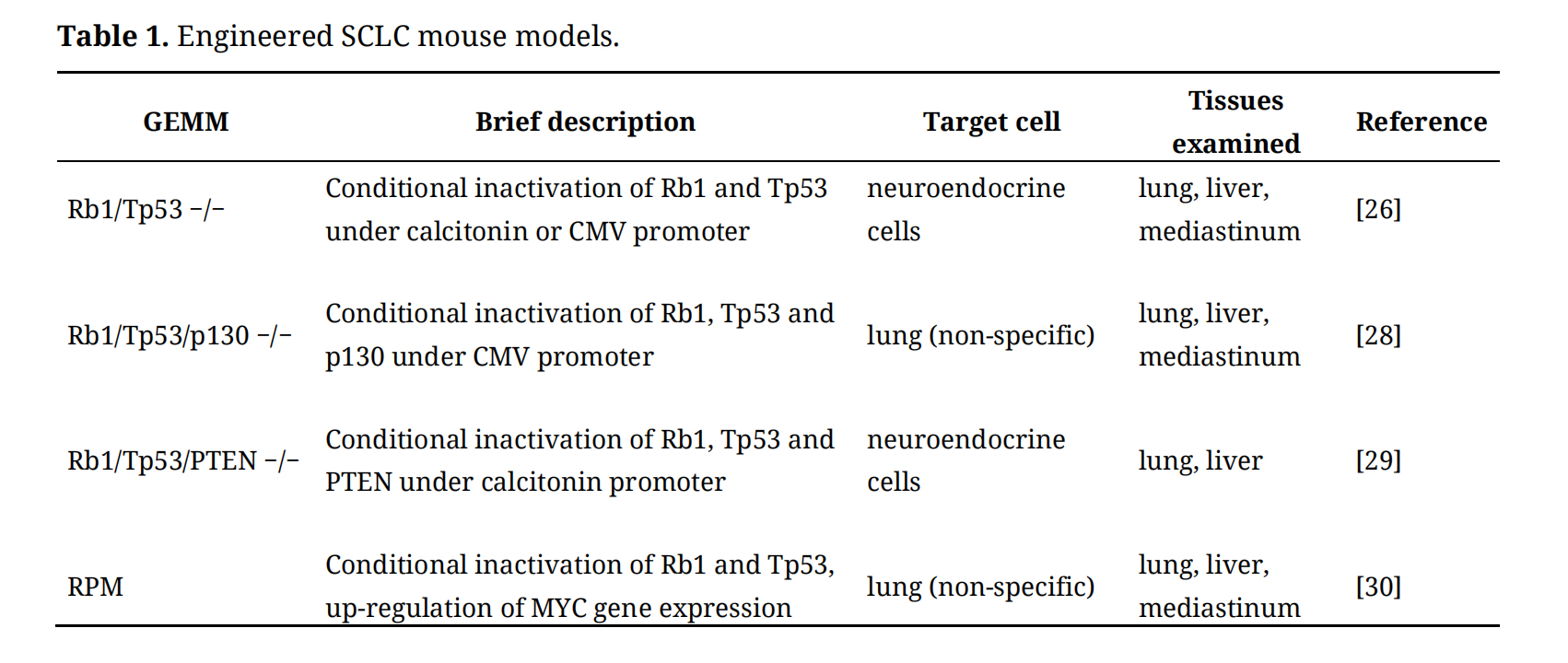 Table 1. Engineered SCLC mouse models.
Table 1. Engineered SCLC mouse models.
The gene expression patterns of both Rb1/Tp53 and Rb1/Tp53/p130 conditional knockout models resemble the ASCL1-high/neuroD1-low subtype of human SCLC [26,28]. Variant Rb1/Tp53/PTEN triple-knockout-mouse model constructed by Jacks Lab is a mixture of large-cell neuroendocrine carcinoma, SCLC and NSCLC [29]. The RPM model is based on the background of Rb1/Tp53 conditional knockout mice with the introduction of a Cre recombinase-activated MYC(T58A) mutant that leads to stabilization of the resultant MYC [30]. The stabilization of MYC strengthened the tumor’s metastatic ability. RPM tumors were mostly central type, mainly in bronchi and bronchioles, growing in a ring shape, and there were two kinds of cells with different morphology. One population, which reflected an ASCL1-high/NeuroD1-low state and appeared often in initial tumor, has the characteristics of typical SCLC, i.e., small cells, less cytoplasm, vague cytoplasm, fine nuclear chromatin particles and inconspicuous nucleolus. The other population, which is often found in invasive tumors and reflected variants of SCLC with ASCL1-low/NeuroD1-high state, is composed of slightly larger cells, single central prominent nucleolus and clear eosinophilic cytoplasm [30].
GEMMs are very useful tools for analyzing the role of individual genes, the interaction between genes and the causes of oncogenes, and they are also helpful when studying drug resistance and developing new treatments. However, GEMMs have many limitations: (1) Human SCLC usually occurs after multiple pathological changes over a long time, while the rapid development of GEMMs often leads to the death of mice before the tumor formation. (2) GEMMs are the only model that can be used to study precancerous lesions and preinvasive lesions of lung cancer. However, the lung cancer in GEMMs is not induced by smoking. Therefore, the genetic changes of the resulting tumors lack the characteristic mutation characteristics of tobacco-related malignant tumors. (3) GEMMs are relatively easy to obtain, and complex models can be quickly constructed by knockout of specific genes. However, the appropriate endpoints of treatment responses, such as overall survival rate and progression-free survival rate, cannot be determined. (4) As a neuroendocrine (NE) cancer, the tumor metastatic pattern in GEMMs is very similar to that of human SCLC. Since SCLC is characterized by early metastatic spread, detailed pathological examination when using GEMMs is necessary.
In recent years, GEMMs have made some great progress. For example, the Rb/Tp53/PTEN triple-knockout-model suggests that ASCL1 plays an important role in the development of SCLC [29]. The inactivation of ASCL1 almost eliminated the formation of neuroendocrine tumors, while the inactivation of NeuroD1 had no significant effect on the number, size and histological appearance of the resulting tumors. However, due to the differences between the targeted mouse gene products and the human corresponding gene products, many drugs that are effective in mouse models have no effect in clinical trials [31,32], which suggests that the main problem in using mouse models to simulate human cancer is that there are inherent biological differences between the two organisms. Therefore, the use of GEMMs to predict clinical reactions and evaluate drug efficacy is debatable.
PDXs are constructed by directly inoculating tumor tissue or cells from a patient’s tumor to immunodeficient mice [33]. They are commonly used to analyze the growth, metastasis and drug sensitivity of human tumors. Compared with GEMMs, PDXs are relatively time-, labor- and cost-effective. Furthermore, PDXs can be used to design personalized targeted therapy [34].
There are two main methods for establishing PDXs: subcutaneous inoculation and intracranial inoculation. Subcutaneous inoculated tumor presents expansive growth, less invasiveness and no metastasis, while intracranial inoculated tumor is highly invasive and lethal. Moreover, intracranial inoculated tumor has a higher success rate of xenograft and requires fewer cells to induce tumor formation [35]. PDXs lay the foundation for acquiring continuous cell lines and tumor tissues for further study.
In recent years, researchers found that circulating tumor cells (CTCs) can be detected in 70%–95% of SCLC patients [36]. New progress has been made in the study of tumor transplantation by transplanting CTCs into mice (CDX model) [37]. Studies have shown that patients with SCLC show a higher CTCs load. CTCs from SCLC patients can effectively form tumors when transplanted into immunodeficient mice [38]. CDX tumor reproduces the histologic characteristics of human SCLC and reflects the effect of chemotherapy. The combined investigation of the preclinical therapeutic study in CDX model with gene profile analysis of human SCLC specimen is a potential and powerful approach to identify chemotherapy-resistant mediators and novel drug candidates. In several clinical trials of SCLC, the CTCs count has been used as an exploratory biomarker of predicting treatment response and drug resistance [39,40]. It can also be used as a pharmacodynamic biomarker in drug development to guide the use of the best therapeutic dose, which has revolutionized the current relatively crude maximum tolerable dose.
PDXs have a wide range of use, which is beneficial to study the tumorigenicity, histological appearance, tumor stem cells and the gene interaction of SCLC. The most common application is in vivo trials of conventional and targeted therapy [41]. Their advantages and disadvantages are also relatively obvious: (1) The histology and gene expression profile of PDX tumor is closer to that of human, and it can better reflect the situation of human SCLC in terms of routine and targeted therapy. However, the tumor inoculated subcutaneously cannot accurately reflect the microenvironment of the tumor. (2) PDXs have advantages for in vitro experiments, drug screening and targeted therapy tests. However, after multiple passages, the mouse matrix gradually replaces the human matrix, which may lead to chaotic results. (3) PDXs are helpful for designing drugs for individualized treatment, but the lack of immune system in the host mice hinders the study of immunotherapy. This obstacle could be overcome by engrafting human CD34+ hematopoietic stem cells. (4) Unlike in humans, diffusion is rare in the PDXs and is genetically difficult to manipulate [42]. (5) Tumors from xenografts could retain their original methylation patterns [43,44], but they may be contaminated by mouse heterosexual viruses.
PDXs were established over 40 years ago [45]. With the development of science and technology, the species of immunodeficient rodent strains have greatly increased, which improves the success rate of xenograft but the cost has also increased greatly. PDXs are often used for the discovery and verification of drugs and biomarkers, and can be used to establish cell lines for biological and other studies. Some PDXs were established in immunocompromised mice, which limits their utility for studies of the interaction between cancer cells and immune cells. The development of humanized xenografts may eventually help resolve this problem [46,47]. Of course, extensive tumor heterogeneity and highly complex mutation spectrum may limit their application value, but PDXs are still the only tool to study complete human tumors in vivo.
In summary, we believe that every model is suitable for a specific research field. For example, PDXs are suitable for timely evaluation of therapeutic responses in vivo, while GEMMs may provide more reliable results in targeted therapy.
With the discovery of the genome and the progress of preclinical models, remarkable progress has been made in understanding the occurrence and development of SCLC and developing new treatments. Different kinds of SCLC preclinical research models are helpful to study the role of various genetic and epigenetic changes in the pathogenesis and biological behavior of SCLC, and are beneficial to study tumor heterogeneity and drug resistance. However, no model can truly summarize all the characteristics of human lung cancer in vivo, and every model has its advantages and disadvantages. It is essential to take full consideration of these advantages and limitations in preclinical trials. At the same time, before selecting the experimental model, the related factors such as cell type, genetic factors, spatiotemporal regulation of target gene expression, tumor microenvironment and metastasis potential of each model should be taken into consideration. Only by comprehensively evaluating these factors, and the application to the proposed research, can we obtain the maximum utility from the preclinical models.
GZ conceived the concept, collected and analyzed references, designed, wrote, and critically revised the manuscript. ZZ and YJ participated in collection and analysis of references, discussions, and partial revision. ZZ also contributed to the critical revision of English language. All authors approved the final manuscript.
The authors have declared no conflicts of interest.
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
41.
42.
43.
44.
45.
46.
47.
Zhao G, Zhao Z, Jia Y. Advances in Preclinical Models of Small Cell Lung Cancer. Med One. 2019;4:e190019. https://doi.org/10.20900/mo.20190019

Copyright © 2020 Hapres Co., Ltd. Privacy Policy | Terms and Conditions




