Location:Home >> Detail
Med One. 2016; 1(1): 4; https://doi.org/10.20900/mo.20160004
 ,
Pei Zhang3
,
Pei Zhang3
1 Department of Endoscopy Center, Shaanxi Provincial People’s Hospital, Xi’an, Shaanxi 710068, P.R. China
2 Department of Digestion Medicine, The Third Affiliated Hospital of Xiangya School of Medicine, Central South University, Changsha, Hunan 410078, P.R. China
3 Department of Radiation Oncology, University of Maryland School of Medicine, Baltimore, MD 21221, USA
*Corresponding Author: Shaojun Liu.
Background: Abnormal p53 expression has been observed in gastric carcinoma cells. The question of whether p53 could become a target for gastric carcinoma biological therapy has not yet been determined.
Methods: TP53 was silenced by siRNA but overexpressed by vector. RT-PCR measured mRNA expression. Western blot measured protein expression. The CCK-8 method was used to analyze cell proliferation. Cell invasion and migration were analyzed, by a Transwell champer and wound healing assay, respectively.
Results: TP53 siRNA significantly silenced TP53 expression and pEGFP-TP53 vector overexpressed p53 protein. Silencing TP53 expression significantly increased cell proliferation, invasion, and migration in BGC-823 cells. In contrast, p53 overexpression significantly inhibited cell proliferation, invasion, and migration in BGC-823 cells.
Conclusion: p53 overexpression may prove a useful strategy for targeted inhibition of gastric carcinoma cell growth and metastasis.
Gastric cancer is one of the most common digestive tract malignant tumors of the digestive tract and it ranks second, in terms of mortality, of all cancers [1-3]. Technological advances in medical imaging technologies and therapeutic advances in chemoradiotherapy, biological target therapy, and cellular immunotherapy have improved gastric carcinoma prognosis. However, it remains poor, which reflects the currently poor understanding of its pathogenic mechanisms [4, 5].
Gastric cancer is a multifactorial disease. Oncogene overexpression and tumor suppressor genes lack of expression are the primary molecular mechanisms involved in tumor growth, therapeutic resistance, and gastric cancer metastasis. Some genes, such as HER-2, VEGFR-2, EGFR, VEGF-A, mTOR, and c-MET/HGF, overexpress has been widely reported in gastric cancer patients and a targeted inhibition of these genes is currently being used as treatment options for gastric cancer [6]. Tumor suppressor genes, such as CDKN2A/p16, CDH1/E-cadherin, RUNX3, and MLH1, are often reported to be inactivated by promoter methylation, mutations, or chromosomal losses [7]. Genomic aberrations in TP53, PIK3CA, ErbB2, ErbB3, ARID1A and KRAS gene are frequently encountered in gastric cancer [8].
TP53 gene mutation is one of the most common anomalies in human cancer. It leads to the loss of TP53 gene expression and p53 functions [9]. TP53 gene mutations are observed in 50-77 % of all advanced gastric cancers [10, 11]. TP53 gene expression downregulation can significantly enhance cell proliferation, migration, and gastric cancer cell invasion [12]. The effects of the p53 protein on gastric cancer cell malignancy are rarely validated by a direct silencing, or an artificially expression, of the p53 protein in gastric carcinoma cell lines.
This study investigated the roles of p53 in the proliferation, invasion, and migration of gastric cancer cells by silencing, and overexpressing, p53 expression in gastric carcinoma BGC-823 cells.
A gastric carcinoma BGC-823 cell line was purchased from the Sunbio Company (Anyang, South Korea). RPMI 1640, and fetal bovine serum, were purchased from Gibco Company (Langley, OK, USA). Transwell chamber and Martrigel were both purchased from BD Bioscience Company (Franklin Lakes, NJ, USA). A lipidosome cell transfection kit was purchased from Invitrogen (Grand Island, NY, USA). Anti-P53 and anti-β-actin antibodies were purchased from Xinlebio (Shanghai, China). A CCK-8 detection kit was purchased from the Beyotime Institute (Shanghai, China). Trizol reagents and a cDNA synthesis kit were purchased from TAKARA (kusatsu, Shiga, Japan). The restriction enzyme and T4 DNA ligase were purchased from NEB (Beijing, China). The plasmid extraction kit was purchased from Omega Bio-tek, Inc (Norcross, GA, USA). The cell protein extraction kit was purchased from Thermo Scientific Company (Waltham, MA, USA).
2.2 SiRNA design and vector constructionThe TP53 siRNA sequence was designed using a publically available online software (www.qiagen.com/siRNA). Complementary siRNA sequences were synthesized and made ready for use by Invitrogen. The TP53 gene was cloned into the pEGFP-N1 vector. cDNA from the TP53 gene was amplified using the forward primer: 5’-CTCGAG GAACAGCTTTGAGGTG-3’ with Xho I sequence in italics and reverse primer: 5'-GGATCC GTCTCTCCCAGGACAGGCACAAAC-3’ with the BamHI sequence in italics. PCR amplification was performed at 94°C, 3min. This was followed by 30 cycles at 94°C, 35s; 58.7°C, 35s; and, 68°C, 1.5 min. Amplified DNA fragments were purified from agarose gel, digested with XhoI and BamH I, ligated into pEGFP-N1 vector (digested with XhoI/BamHI), and transformed into DH5α competent cells. Clones were selected and verified by sequencing. The vector produced was designated as “pEGFP-TP53”.
2.3 Cell culture and transfectionBGC-823 cells were cultured in an RPMI 1640 medium containing 10 % fetal bovine serum. The cells were transformed with siRNA or pEGFP-TP53 plasmid using a lipidosome cell transfection kit and following the instructions contained in the user mannual.
2.4 RNA extraction and reverse transcriptionCells were harvested and centrifuged. Cell pellets were used for RNA isolation with TRizol reagent by following the instructions contained in the manufacturer’s manual. 1 ug of RNA was used for cDNA reverse transcription using a cDNA synthesis kit. Synthesized cDNA was kept at -20°C.
2.5 Western blotTotal protein was extracted from cultured cells using a protein extraction kit and following the instructions in the manufacturer's manual. Protein concentration was determined by a BCA protein assay. Protein aliquots were kept at -80°C. Western blot was performed as described above [13]. Forty µg of protein was loaded for each lane and separated on 10 % SDS-PAGE gels. After transferring and blocking, the membranes were incubated with either anti-p53, or β-actin, antibody overnight at 4°C. This was followed by a secondary antibody for 1 hr at room temperature. Bands were visualized using ECl reagents and scanned for density analysis. β-actin expression was used as an internal control for sample loading.
2.6 Cell proliferation assayCells in 96-well plates were transfected with siRNA, or plasmid, and continuously incubated for 6 hrs, 12 hrs, 24 hrs, and 36 hrs. Ten µl of CCK8 solution was added to each well and the cells were continuously incubated for 3 hrs. Absorbance values were read at 450 nm in an ultraviolet spectrophotometer.
2.7 Cell invasion assayInvasion assay was conducted in a 24-well Transwell chamber with matrigel-coated membranes. Two hundred ul of (1 × 105 cells/ml) cell suspensions in a serum free medium were spread onto the wells in the upper chamber and 1000 ul of complete cell medium containing 10 % FBS were applied to the wells in the lower chamber. Incubation for 12, 24, and 36 hrs at 37°C, 5 % CO2 followed. Membranes in the lower chamber were removed and stained with 0.1 % crystal violet dye for 15 min. The cells were counted under a Fluorescence Inversion Microscope.
2.8 The cell migration assayCell migration was analyzed using a wound healing assay. Seeded into 6-well plates were 5 × 105 cells. After transfection, cells were cultured for 24h and then a scratch line was made using pipette tips. After rinsing three times with 1 × PBS, cells were continuously cultured in a serum-free medium for 24 hrs. After photographing, scratch width in the photographs was analyzed using Image J software.
2.9 Data analysisData were analyzed using SPSS software and presented as mean ± standard error. Variance Analysis was used for statistical difference. A p < 0.05 was considered statistically different.
TP53 siRNA transfection was significantly downregulated (Fig. 1A). pEGFP-TP53 plasmid transfection significantly increased (Fig. 1B) TP53 mRNA expression. pEGFP-TP53 transfection resulted in a 278 % increase in p53 protein expression (Fig. 1C, 1D). TP53 siRNA transfection resulted in a 67 % decrease in p53 protein expression (Fig. 1C, 1D). These findings suggest that the TP53 gene was successfully both silenced and overexpressed.
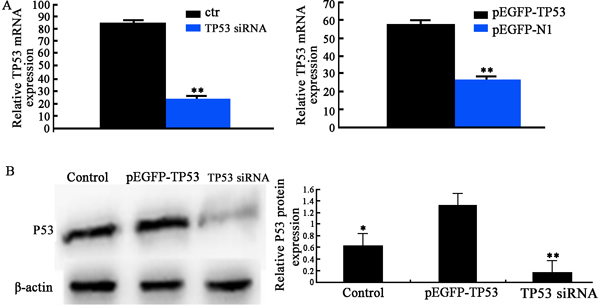 Fig. 1 TP53 gene expression.
Fig. 1 TP53 gene expression.
A) RT-PCR detection of TP53 mRNA expression after transfection of TP53 siRNA (left panel) and pEGFP-TP53 plasmid (right panel). **p < 0.01 between two groups. B) Western blot of P53 protein expression. Left panel: representative Western blots. Right panel: semi-quantatitive analysis of Western blots. *p < 0.05, **p < 0.01 vs. pEGFP-TP53.
Cell proliferation was analyzed using a CCK-8 kit. TP53 siRNA transfection significantly increased proliferation in BGC-823 cells (p < 0.05). pGEFP-TP53 plasmid transfection significantly decreased proliferation in BGC-823 cells (p < 0.05) (Fig. 2).
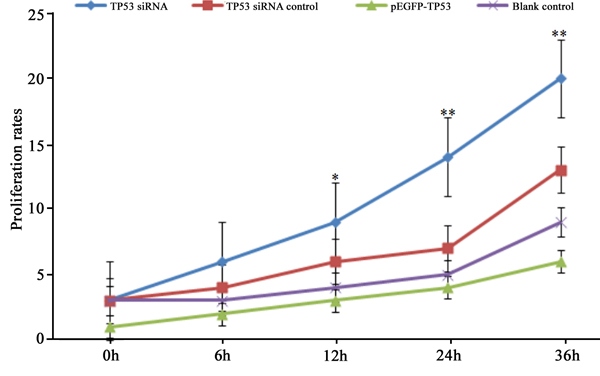 Fig. 2 Cell proliferation rates.
Fig. 2 Cell proliferation rates.
Cells were transfected with TP53 siRNA or pEGFP-TP53 plasmid. *p < 0.05, **p < 0.01 vs. pEGFP-TP53
A wound-healing assay showed that TP53 siRNA transfection significantly increased cell migration (p < 0.05) and that pGEFP-TP53 plasmid transfection significantly decreased cell migration in BGC-823 cells (p < 0.05) (Fig. 3).
 Fig. 3 The effect of p53 on cell migration.
Fig. 3 The effect of p53 on cell migration.
Migration was analyzed by wound healing assay.
The Transwell invasion assay showed that TP53 siRNA transfection significantly increased cell invasion (p < 0.05), but that pGEFP-TP53 plasmid transfection significantly decreased cell invasion in BGC-823 cells in a time-dependent manner (p < 0.05) (Fig. 4).
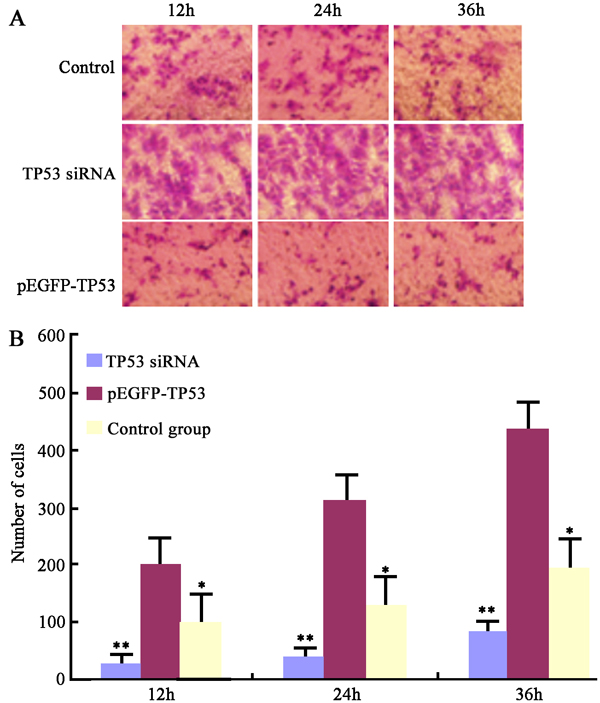 Fig. 4 The effect of p53 on cells invasion.
Fig. 4 The effect of p53 on cells invasion.
A) Images of transwell champer assay. B) Cell counting of transwell chanper assay. *p < 0.05, **p < 0.01 vs. TP53 siRNA transfection at the same time point.
Despite an increasing comprehension of molecular mechanisms and the consequent development of targeting therapy for gastric cancers, the prognosis remains poor, particularly in advanced disease patients. TP53 gene mutations have been reported as present in as many as 77 % of metastatic tumor patients [11]. These mutations impair p53 tumor suppressor effects, such as inhibiting proliferation, migration, and invasion of gastric cancer cells [12]. There is no direct evidence as to whether restoring TP53 gene function could inhibit gastric tumor growth and metastasis. This study detected that silencing of TP53 gene expression significantly increases cell proliferation, invasion, and migration in gastric carcinoma cells, while p53 overexpression significantly inhibits cell proliferation, invasion, and migration.
The functions of the p53 protein includes controlling the cell cycle, inhibiting proliferation, inducing cell apoptosis, stimulating functional factor synthesis, and regulating cell metabolism. It also takes part in DNA damage repair [14]. p53 dysfunction is also an important factor driving gastric cancer carcinogenesis and progression. Various types of TP53 gene mutations not only damage its tumor suppressor function, but also produces new functions that promote tumor development and progression. In this study, silencing TP53 gene expression enhanced the malignant phenotype of human gastric adenocarcinoma cells. In contrast, expression of a wild-type p53 protein restored p53 tumor suppressive functions in gastric adenocarcinoma cells. This study indicates that p53 plays a very important role in regulating the proliferation, migration, and invasion of gastric carcinoma cells. Restoring TP53 gene expression can restore its p53 tumor suppressive functions and can be used as a treatment strategy.
In conclusion, TP53 overexpression provides an attractive strategy for targetting inhibition of gastric carcinoma cell growth and metastasis.
The authors declare that they have no conflict of interests.
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
Li Y, Liu S, Zhang P. Direct Evidence for TP53 Involvement of TP53 in Proliferation, Migration, and Invasion of Gastric Carcinoma Cells. Med One. 2016; 1(1): 4; https://doi.org/10.20900/mo.20160004

Copyright © 2020 Hapres Co., Ltd. Privacy Policy | Terms and Conditions



